|
Spis treści:
Zasady i sposoby pobierania próbek paleontologicznych - JERZY DZIK
Geologia i paleontologia Dzikowca
- BŁAŻEJ BERKOWSKI
Flora i środowisko karbonu produktywnego Nowej Rudy - SŁAWOMIR FLORJAN
Roślinność karbonu na tle flor paleozoiku - GRZEGORZ PACYNA
Flora mezozoiku jako tło dla roślin kopalnych z Krasiejowa - ANNA MARIA OCIEPA
Flora kajpru - DANUTA ZDEBSKA
Małże kajpru Krasiejowa - MICHAŁ BRODACKI
Znaczenie okresu triasowego w ewolucji ryb - ŁUKASZ PAŚKO
Kapitosaury - DOROTA MAJER
Metopozaury - TOMASZ SULEJ
Fitozaury - MARIUSZ LUBKA
Aetozaury - TOMASZ SULEJ
Dinozaury triasu - JERZY DZIK
Geneza dwunożności dinozaurów - EWA BARYCKA
Zasady i sposoby pobierania próbek paleontologicznych
JERZY DZIK
Instytut Paleobiologii PAN
I szkielet dinozaura jest próbką paleontologiczną. Próbką, w której jest tylko jeden (duży) okaz skamieniałości. Do pewnych celów badawczych to już dość, do innych nie. Anatomowi jeden okaz dostarczyć może materiału na całą karierę naukową; badaczowi stosunków ekologicznych kilkaset okazów da materiał na niezły artykuł naukowy; dla zainteresowanego przemianami ewolucji na poziomie populacji i sto tysięcy skamieniałości to za mało. Pobierając próbki paleontologiczne trzeba więc mieć na względzie cel, któremu ma to służyć.
Po co?
Dane paleontologiczne mają dwa główne zastosowania naukowe:
(1) Służą do odtwarzania sytuacji środowiskowej w dawnych epokach geologicznych. Są uzupełnieniem i pozwalają na sprecyzowanie wyników badań sedymentologicznych. Pozwalają również na prześledzenie zmian środowiska w czasie, co jest podstawą wydzielenia jednostek skalnych w biostratygrafii i szczegółowej korelacji wiekowej.
(2) Pozwalają na odtworzenie przemian ewolucyjnych organizmów. Bez tego nie do pomyślenia jest odtworzenie przebiegu filogenezy, ani wydzielanie jednostek czasu geologicznego.
Wypełnianie obu tych zadań wymaga opróbkowania ze stosowną precyzją. Interpretacja pozyskanych danych wymaga zaś zrozumienia mechanizmów powstawania skamieniałości i biologicznego sensu ich nagromadzeń w skałach.
Odtwarzanie środowiska.- Opróbkowanie w paleontologii ma nieco odmienny sens, niż w biologii. Odmienność tę najprościej uzmysławia różnica zawartości dostępnych informacji pomiędzy próbką szkieletów wziętą bezpośrednio z żywej populacji (np. w wyniku kataklizmu takiego, jaki przytrafił się niegdyś Pompejom), a próbką ze zwykłego cmentarza. O ile w materiale z Pompei zapisana jest struktura rozkładu biomasy w obrębie populacji, to skład szkieletów na cmentarzu odzwierciedla specyfikę produkcji biologicznej. Odnosi się to zarówno do struktury wiekowej próbki, jak i do składu gatunkowego całej biocenozy. W żywej populacji dominują stadia wzrostowe, których jest najwięcej w danym momencie; na cmentarzu zaś te, których dotyczy najwyższa śmiertelność. Określenie dynamiki populacji (rozkładu śmiertelności) w każdym z tych przypadków wymaga odmiennych operacji rachunkowych. Oznacza to, że w trakcie pobierania próbek paleontologicznych trzeba próbować ocenić, z którym rodzajem zespołu kopalnego ma się do czynienia: efektem katastrofy, czy zwykłym cmentarzyskiem. Dobrze więc wiedzieć, co nieco o sedymentologii, nawet jeśli się jest ściśle biologicznie orientowanym badaczem kości czy liści. Okresowo zrzucane liście dają bardzo szczególny rodzaj zespołu skamieniałości. Jego istote w drastyczny sposób ukazują próbki zawierające wylinki stawonogów (np. trylobitów czy małżoraczków). Mimo, iż są to "próbki z cmentarza", ich struktura wiekowa jest taka, jak skład "próbek z biomasy".
Poznanie przebiegu ewolucji.- Paleontologia dostarcza danych pozwalających na odtworzenie przebiegu ewolucji na dwu skrajnych poziomach. Z jednej strony są to znaleziska wiążących ogniw ewolucji i tu trudno zwykle liczyć na więcej, niż pojedyncze okazy - fragmenty szkieletów czy organów roślinnych. Geologiczna pozycja takich znalezisk, zarówno od strony stratygrafii, jak środowiska sedymentacji, jest równie ważna, co informacje morfologiczne. Znaleziska takie wymagają więc nie mniej solidnej dokumentacji, co inne próbki paleontologiczne. Drugim skrajem różnorodności zapisu kopalnego są znaleziska umożliwiające badania przemian ewolucyjnych na poziomie populacji. Tu liczy się precyzja w pobieraniu próbek i liczebność okazów. Ile okazów zbierzemy, to ograniczenie techniczne, wynikające z kosztów i czasu badań. Są jednak i bardziej zasadnicze powody, dla których nie warto mnożyć, ani nazbyt zagęszczać pobierania próbek.
Jak gęsto?
Uśrednienie czasowe (time-averaging) czyni próbki paleontologiczne czymś fundamentalnie odmiennym od próbek pobieranych przez biologów. Uniemożliwia, rzecz jasna, rozważanie zjawisk dziejących się w krótkich odcinkach czasu (godzin, dni, tygodni, czy miesięcy). Z drugiej jednak strony usuwa z zapisu kopalnego (odfiltrowuje) znaczna część nieistotnych fluktuacji, wzmacniając zjawiska trwałe. Zapis kopalny jest przez to od razu, bez dodatkowej obróbki statystycznej, bliższy głębszej istocie zjawisk ekologicznych czy ewolucyjnych od materiału zbieranego przez biologów. Dane eksperymentalne pozwalają na oszacowanie reprezentatywności zapisu kopalnego. Jest ona różna, zależnie od środowiska. Są sytuacje, kiedy osiąga ścisłość niedostępną nawet badaczom dzisiejszych organizmów.
Pobierając próbki warto jednak mieć na względzie ograniczenia wynikające z samej natury zapisu kopalnego. W przypadku opróbkowywania w celu pozyskania materiałów mikroskopijnych skamieniałości dwa czynniki są szczególnie istotne. To tempo formowania osadu i głębokość jego homogenizacji przez ryjące organizmy. Jak istotne, można ocenić na podstawie charakteru skały.
Tempo sedymentacji.- Jest wyraźna zależność tempa formowania osadu od jego pochodzenia i wielkości ziarna. Z reguły formacje piaskowcowe czy zlepieńcowe powstawały bardzo szybko i setki metrów piaskowca bywają wiekowym odpowiednikiem decymetrów wapieni. Stosowna do tego musi więc być i gęstość opróbkowania. Nierzadko bywa tak, że większość czasu geologicznego nie ma w ogóle zapisu w skale. Czas ukrywa się wtedy raczej w fugach między warstwami. Wynika to zwykle z usunięcia części wcześniej złożonego osadu przez erozję poprzedzającą kolejny epizod sedymentacji. Im dalej od głębokomorskich środowisk oceanicznych, tym więcej takich luk stratygraficznych. W środowiskach kontynentalnych, gdzie erozja dominuje, to raczej samo uformowanie się formacji skalnej jest ewenementem.
Wniosek z tego taki, że istotniejsze jest pobranie próbek z granic między jednostkami stratygraficznymi, czy wręcz warstwami (jedna próbka nad granicą, druga pod nią), niż opróbkowywanie wewnątrz samej jednostki. Z drugiej strony powolna i jednostajna sedymentacja wewnątrz dłużej trwających epizodów formowania osadu dostarczyć może najlepszego, pełnego zapisu ewolucji. Ewolucji biologicznej i ewolucji środowiska. W przypadku ewolucji środowiska opróbkowanie powinno być odpowiednio gęste, by nie przeoczyć żadnej zmiany. Badaczom ewolucji biologicznej wystarczy, że populacje reprezentowane przez próbki sąsiadujące ze sobą nie różnią się istotnie od siebie.
Jeśli pobieranie próbek zostanie odpowiednio dobrze udokumentowane, zawsze można sprawdzić, czy między podejrzanymi próbkami nic istotnego się nie zdarzyło wróciwszy do odsłonięcia. Jeśli odsłonięcie nie jest czynnym kamieniołomem, dobrze jest pozostawić namalowane sprayem numery pobranych próbek na ścianie skalnej. Nie będzie wówczas problemów z identyfikacją miejsc pobrania próbek i nie potrzeba od razu przeprowadzać pełnego opróbkowania. Z czynnych kamieniołomów roztropnie wziąć więcej skały, niż potrzeba. Łatwiej wyrzucić próbkę, niż uzupełnić opróbkowanie. Są jednak obiektywne granice opłacalnej gęstości opróbkowania.
Bioturbacja.- Uśrednienie czasowe jest właściwością wszelkiego materiału kopalnego. Jest wynikiem nie tylko fizycznej natury sedymentacji, związanej z przemieszczaniem i przemywaniem osadu. Osad jest też mieszany przez organizmy, czyli bioturbowany. Głębokość, do której dokonuje się całkowite zhomogenizowanie osadu jest granicą, poza którą nie ma sensu zagęszczanie próbek - wszystkie będą bowiem miały ten sam skład. Głębokość bioturbacji jest odmienna w różnych środowiskach. Nie ma jej w osadach pozbawionych tlenu, bo nie mogą w nich żyć organizmy ryjące. Skały nie zbioturbowane mają na przełamie horyzontalną laminację (pamiętać jednak należy, że podobna laminacja może być wynikiem kompakcji, czyli sprasowania osadu ilastego przez jego odwodnienie). Głębokość bioturbacji w poszczególnych rodzajach środowisk podlegała też zmianom w trakcie ewolucji biologicznej organizmów ryjących. We wczesnym paleozoiku była płytsza (zwykle kilka centymetrów), niż w mezozoiku i dziś (często ponad metr). Dobry zapis kopalnych wymaga więc skał stosunkowo słabo bioturbowanych.
Jak dużo?
Dobrze jest mieć jak najobfitszy materiał. Ograniczenia stawia jednak przyroda, limitując jego dostępność, a także zdrowy rozsądek wsparty przez zwierzchników, pilnujących terminów przygotowywania dysertacji i publikacji. Statystyka dyskwalifikuje próbki zawierające mniej, niż dwadzieścia okazów jako źródło informacji o strukturze gatunkowej zespołu czy strukturze wiekowej populacji. Trzeba się więc starać, by w próbce znalazło się kilkaset okazów nadających się do analizy. W przypadku badań ewolucji na poziomie populacji oznacza to konieczność pobierania ogromnych próbek, bo przecież badane gatunki stanowią często ledwie kilka procent składu całego zespołu kopalnego. W nauce dzisiejszej trzeba się jednak dobrze napracować, by osiągnąć publikowalne wyniki, i nie ma na to rady. Naszym następcom będzie jeszcze trudniej.
Również biostratygrafia nakazuje posługiwać się próbkami o rozmiarach nadających informacji o morfologii organizmów sens statystyczny. Zmienność wewnątrz populacji ma gaussowski rozkład, co oznacza, że im mniej typowa dla populacji cecha, tym mniejsze prawdopodobieństwo jej wystąpienia w populacji. W praktyce do określenia z dużym prawdopodobieństwem, które cechy są właściwe dla populacji wystarcza kilka okazów. Bardzo ryzykowne jest jednak wnioskowanie o wieku warstw skalnych na podstawie pojedynczych okazów. Pozbawione zaś sensu jest z tego względu posługiwanie się tzw. wertykalną (czyli typologiczną) koncepcją gatunku. Zasięg gatunków przewodnich, a przez to i jednostek biostratygraficznych, uzależniony jest wówczas bezpośrednio od wielkości próbki.
Rady praktyczne
Po prawidłowo przeprowadzonym opróbkowanie pozostać powinna pełna dokumentacja muzealna. Przede wszystkim same próbki paleontologiczne, czyli wypreparowane makroskamieniałości w pudełkach z etykietkami i numerami na okazach, opis i rysunek profilu geologicznego oraz próbki litologiczne. Dobrze wykonywać dokumentację według ustalonego, możliwie powszechnego standardu. Poniżej przedstawiam sugestie, jak taki standard mógłby się przedstawiać w odniesieniu do materiałów paleontologicznych zebranych z terytorium Polski.
Rysunek profilu.- Od tej wstępnej czynności zależy sens dalszego postępowania. Profil geologiczny przedstawiany jest zwykle w postaci słupka 2-3 cm szerokości z możliwie wiernie odtworzonym następstwem warstw. Jeśli poszczególne warstewki skalne są od siebie wyraźnie odmienne, oznacza się ich naturę odpowiednimi symbolami (szrafurą: piasek kropkami, wapień kostkami, ił poziomymi kreskami, itp.). Prawa strona słupka jest nierówna, co pozwala na zaznaczenie fizycznych właściwości warstw skalnych. Im bardziej wystają poza linię pionową, tym grubsze ziarno, lub (co łatwiej ocenić w terenie bez badań mikroskopowych) większa odporność na wietrzenie. Profil należy rysować osobiście na papierze milimetrowym, nawet jeśli był już wcześniej przez kogoś zmierzony i opublikowany. Nie trzeba się sugerować cudzymi wynikami - dopiero po sporządzeniu rysunku warto porównać własne dokonania z opublikowanymi. Skala rysunku powinna być mała, najlepiej 1:10; jeśli profil jest dłuższy - 1:20. Powinny się na nim znaleźć wszystkie warstewki skalne rozpoznawalne na podstawie litologii czy barwy. Większe pomniejszenia robi się już kserograficznie z oryginalnego rysunku.
Położenie próbek.- Nanosi się je na rysunek profilu z maksymalną dokładnością tak, by nie mieć później wątpliwości, z której warstewki próbka pochodzi. Nie ma znaczenia kolejność numeracji, pod warunkiem, że nie oznaczy się tym samym numerem odmiennych próbek. Dlatego każdy profil geologiczny powinien mieć swoje osobne oznaczenie literowe albo cyfrowe, które jest częścią numeru próbki. Wszystkie próbki, nawet te wzięte z rumoszu, powinny być wpisane na rysunku (choćby na jego marginesie), by nie zapomnieć, czy jakiegoś numeru już się nie użyło.
Opakowanie próbki.- By uniknąć efektu młyna kulowego i kompletnego roztarcia skały, dogodnie jest brać próbkę w postaci jednego bloku skalnego. Jeśli próbka zawiera duże skamieniałości, każdą z nich wypada osobno owinąć w miękki papier. Do opakowania całej próbki dobre są grube (dwuwarstwowe) papierowe torby, na których wypisuje się numer. Papier wchłania i przepuszcza wilgoć, więc próbki szybko wysychając nie pleśnieją, a napis na torbie nie niszczeje. Torby z tkaniny są drogie; polietylenowe torebki zaś niepraktyczne, bo trudno je opisać, rozrywają je ostre krawędzie skalne i trzymają do tego wilgoć.
Próbki litologiczne.- Po każdej próbce dobrze jest pozostawić ułamek skały. W przyszłości można go wykorzystać do wykonania płytki cienkiej (szlifu), badań izotopowych czy geochemicznych. Szczególnie wygodne jest przygotowanie próbek litologicznych w postaci wyciętych tarczą diamentową płytek o grubości około 3 mm i rozmiarach 3×7 cm. Można je ustawiać pionowo w pudełku, a numer próbki wypisany czarnym tuszem w górnym rogu ułatwia przeszukiwanie.
Preparacja skamieniałości.- Przenoszenie próbek do pojemników i wypisywanie etykiet jest częstym źródłem pomyłek, dobrze jest więc zachować fragment oryginalnej torby z numerem i użyć go jako etykietkę, czy jej duplikat. W przypadku próbek zawierających duże skamieniałości koniecznie należy po umyciu skamieniałości wypisać na nich trwałym tuszem numer próbki lub choć odsłonięcia. Uniknie się wtedy "odkryć" naukowych wynikłych z przypadkowego przełożenia skamieniałości do niewłaściwego pudełka. Na skamieniałościach przeznaczonych do naukowego opracowania powinien być numer kolekcji muzealnej. Materiały takie muszą być bowiem zdeponowane w publicznej instytucji muzealnej. Wygodnym i prostym sposobem nanoszenia numerów jest wypisanie ich tuszem na maleńkim papierowym krążku z dziurkacza i przyklejenie do skamieniałości. Krążek taki jest dobrze widoczny i łatwy do usunięcia w razie potrzeby.
Literatura
- Dzik, J. 1996. Dzieje życia na Ziemi. Wprowadzenie do paleobiologii. Wydanie 2., zmienione i poprawione. 515 pp. Państwowe Wydawnictwo Naukowe. Warszawa. (wydanie 3. w przygotowaniu)
Zawiera objaśnienie zagadnień reprezentatywności zapisu kopalnego.
- Goldring, R. 1999. Field Palaeontology. 2nd Edition. 208 pp. Longman, London.
Doskonały podręcznik sedymentologicznie orientowanej paleontologii z wprowadzeniem do tafonomii i ichnologii.
- Kowalewski, M. 1996. Time-averaging, overcompleteness, and the geological record. Journal of Geology 104, 17-326.
- Kowalewski, M., Goodfriend, G.A., Flessa. K.W.1998. High-resolution estimates of temporal mixing within shell beds: the evils and virtues of time-averaging. Paleobiology 24, 287-304.
To eksperymentalne prace dowodzące, że w przypadku muszli małżów uśrednienie czasowe pozwala na rozpatrywanie jedynie zjawisk w stuletnich czy tysiącletnich skalach czasu. Wówczas zapis jest reprezentatywny. Rezultatem uśrednienie czasowego jest swoista nadreprezentatywność: w zapisie kopalnym współwystępują ze sobą nawet organizmy normalnie się ze sobą nie spotykające.
- McKinney, M. 1985. Distinguishing patterns of evolution from patterns of deposition. Journal of Paleontology 59, 561-567.
Analiza związku pomiędzy kompletnością zapisu geologicznego a środowiskiem sedymentacji, pomocna przy określaniu gęstości opróbkowania.
- Meldahl, K.H., Scott, D., & Carne, K. 1995. Autochthonous leaf assemblages as records of deciduous forest communities: An actualistic study. Lethaia 28, 383-394.
Eksperymentalna praca dowodząca, tym razem na przykładzie liści drzew, że uśrednienie czasowe zapisu kopalnego nie daje szans na zapis krótkotrwałych zjawisk. Zapis jest jednak reprezentatywny dla długotrwałej sytuacji zbiorowiska roślinnego.
Raup, D.M. & Stanley, S.M. 1984. Podstawy paleontologii. 525 pp, PWN, Warszawa.
Wciąż aktualny podręcznik metod wnioskowania w paleontologii, w tym również opis sposobów wykorzystania próbek.
- Thayer, C.W. 1983. Sediment-mediated biological disturbance and the evolution of marine benthos. In; - - M.J.S. tevesz & P.L. McCall (eds) Biotic Interactions in Recent and Fossil Communities. 479-625. Plenum Press, New York.
Dyskusja o "teorii buldożera", zgodnie z którą w trakcie fanerozoiku zwiększała się głębokość bioturbacji - istotne przy określaniu gęstości opróbkowania.
- Vermoere, M., Vanhecke, L., Waelkens, M., & Smets, E. 2000. A comparison between modern pollen spectra of moss cushions and Cundill pollen traps - Implications for the interpretation of fossil pollen data from Southwest Turkey. Grana 39, 146-158.
Kolejna analiza reprezentatywności zapisu pyłkowego. Standardowe pułapki pyłku porównane z poduszkami mchów dokumentujacymi wieloletnie skumulowanie zestawu pyłku.
Geologia i paleontologia Dzikowca
BŁAŻEJ BERKOWSKI
Uniwersytet Poznański
Kamieniołom Wapnica w Dzikowcu położony jest w północnej części tektonicznej struktury bardzkiej. Od południa oddzielony jest od masywu gabrowo-diabazowego Nowej Rudy-Słupca przez tektoniczny rów Czerwieńczyc. Sam kamieniołom jest od kilkudziesięciu lat nieczynny, stąd prześledzenie całego profilu dewońskich i karbońskich utworów jest obecnie niemożliwe. Do uzupełniania opisu mogą jednak posłużyć dane badaczy niemieckich, w szczególności Güricha (1902), który sporządził pięć kolejnych przekrojów litologicznych W-E od południa na północ. Gabro stanowiące część masywu gabrowo-diabazowego Nowej Rudy tworzy krystaliczną podstawę profilu. Sekwencję rozpoczynają warstwy podstawowe (ok. 15 m) zalegające na gabrze, opisywane przez Güricha (1902) jako wapienie z otoczakami gabra (a), na których kolejno zalegają: wapień krynoidowy (b), wapień jasno-brązowy (g) i czerwony wapień skorupowy z miałem gabrowym (d).
Na podstawie skamieniałości znalezionych w wypełnieniach pomiędzy blokami gabra przyjąć można co najmniej wczesnofameński (>370 Ma) wiek gabra. Ostatnio przeprowadzone datowania radiometryczne metodą Sm-Nd (Pin et al. 1988) określiły wiek masywu na 351 ą 21 Ma (mln lat). Datowanie paleontologiczne z pewnym trudem mieści się więc tu w granicach błędu oznaczenia radiometrycznego.
Nad sekwencją wapienia podstawowego znajduje się ok. 40 m wapienia głównego (Tietze 1870), opisanego przez Güricha (1902) jako wapień gruzłowy oznaczony symbolem (e). Wapień ten był określany przez badaczy niemieckich jako ubogi w skamieniałości wapień frański. Za niekwestionowany famen uważano wtedy tylko wapień klymeniowy (Tietze 1870; Gürich 1902; Bederke 1924), ok. 3 m miąższości. Z wapienia tego pochodzą pierwsze dewońskie amonity znalezione na obecnym terytorium Polski (Buch 1838). Schindewolf (1937; także Becker 2000) dokładnie oznaczył głowonogi z wapienia klymeniowego i wyróżnił w nim niższą część należącą do późnofameńskiego piętra Wocklumeria i wyższą do wczesnoturnejskiego piętra Gattendorfia oraz wykazał istnienie luki stratygraficznej pomiędzy tymi piętrami (powyżej poziomu Parawocklumeria paradoxa). Szczegółowe badania stratygraficzne piętra Gattendorfia oparte na głowonogach i konodontach przeprowadził Dzik (1997). Cały profil węglanowego famenu i najniższego turneju od podstawy krystalicznej do niezgodnie leżącego na nim okruchowego turneju bądź wizenu (kulmu) ma ok. 60 m miąższości.
Szczegółowy opis odsłonięć zamieszczony został w powielonym przewodniku Kursu.
Literatura
Becker, R.T. 2000. Taxonomy, evolutionary history and distribution of the middle to late Famennian Wocklumeriina (Ammonoidea, Clymeniida). Mitteilungen der Museum für Naturkunde in Berlin, geowisseschaftliche Reihe 3, 27-75.
Bederke, E. 1924. Das Devon in Schlesien und das Alter des Sudetenfaltung. Forschritte der Geologie und Paleontologie 2, 1-50.
Buch, L. von 1838. Über Goniatiten und Clymenien in Schlesien. Physikalische Abhandlungen der königliche Akademie der Wissenschaften in Berlin 1838, 149-169.
Chorowska, M. & Radlicz, K. 1984. Karbońskie wapienie i zlepieńce zachodniej części struktury bardzkiej. - Kwartalnik Geologiczny 28, 251-290.
Chorowska, M. & Radlicz, K. 1987. Wapienie górnego dewonu i dolnego karbonu w kamieniołomie na górze Wapnica w Dzikowcu. Przewodnik 58 Zjazdu Polskiego Towarzystwa Geologicznego, Wałbrzych.
Dzik, J. 1997. Emergence and succession of Carboniferous conodont and ammonoid communities in the Polish part of the Variscan sea. Acta Palaeontologica Polonica 42, 57-170.
Gürich, G. 1902. Zur Discussion über das Profil von Ebersdorf. Zeitschrift der Deutschen Geologischen Geselschaft 54, 57-65.
Haydukiewicz, J. 1990. Stratigraphy of Paleozoic rocks of the Góry Bardzkie and some remarks on their sedimentation (Poland). Neues Jahrbuch für Geologie und Palaontologie, Abhandlungen 179, 275-284.
Lewowicki, S., 1959. Fauna wapieni klimeniowych z Dzikowca Kłodzkiego. Biuletyn Instytutu Geologicznego 146, 73-118.
Oberc, J. 1972. Sudety i obszary przyległe. Budowa geologiczna Polski Tom 4. Tektonika, część 2. 307 pp. Wydawnictwa Geologiczne, Warszawa.
Pin, C., Majerowicz, A., & Wojciechowska, I. 1988. Upper Palaeozoic oceanic crust in the Polish Sudetes: Nd-Sr isotope and trace element evidence. Lithos 21, 195-209.
Schindewolf, O.H. 1937. Zur stratigrafie und Palaeontologie der Wocklumer Schichten (Oberdevon). Preussische Geologishe Landesanstalt Abhandlungen Neue Folge 178, 1-132.
Tietze, E. 1870. Über die devonischen Schichten von Ebersdorf umweit Neurode in der Grafschaft Glatz, eine geognostisch-paläontologische Monografie. Palaeontographica 19, 108-158.
Wajsprych, B. 1986. Sedimentary record of tectonic activity on Devonian-Carboniferous continental margin, Sudetes. IAS 7th European Regional meeting. Excursion Guidebook. Kraków-Poland 141-164.
Flora i środowisko karbonu produktywnego Nowej Rudy
SŁAWOMIR FLORJAN
Uniwersytet Jagielloński
Nowa Ruda położona jest na terenie Dolnośląskiego Zagłębia Węglowego, którego przedłużenie znajduje się po stronie czeskiej. Jest to najmniejsze z trzech naszych zagłębi węgla kamiennego i jednocześnie jedyne, w którym zakończono już eksploatację węgla. W rejonie Wałbrzycha była ona prowadzona przez kopalnie "Thorez", "Wałbrzych"" i "Victoria", a w rejonie Nowej Rudy przez kopalnię "Nowa Ruda". Obecnie okazy flory karbońskiej można pozyskiwać w Zagłębiu jedynie z dostępnych jeszcze hałd kopalnianych (co ma niewielką wartość stratygraficzną) oraz z odsłonięć terenowych.
Zagłębie Dolnośląskie leży w depresji śródsudeckiej. Jest ono zagłębiem limnicznym, typu orogenicznego, które powstało w zapadlisku śródgórskim. Utwory późnego karbonu mają miąższość około 1700 m. W ich obrębie występuje około 80 pokładów węgla o miąższości do 3 m (Gabzdyl 1994; Bossowski et al. 1995).
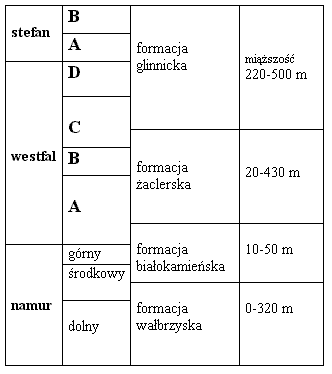 | | Ryc. 1. Podział stratygraficzny utworów późnego karbonu rejonu Nowej Rudy (wg Bossowskiego et al. 1995). |
W późnym karbonie w rejonie Nowej Rudy (Ryc. 1) dominowały rzeczne środowiska sedymentacyjne. Z górnej części profilu utworów górnego karbonu znane są też osady środowisk jeziornych. Zasoby bilansowe węgla kamiennego znajdują się w utworach od wczesnego namuru do westfalu B, a więc w formacjach wałbrzyskiej i żaclerskiej (Ryc. 1). Syntetyczne opracowanie zagadnień dotyczących geologii utworów karbonu w depresji sródsudeckiej jak i na całym Dolnym śląsku wraz z bibliografią znaleźć można np. w pracach pod redakcją Bojkowskiego i Porzyckiego (1983) oraz Zdanowskiego i Żakowej (1995).
Makroflora karbońska znana jest z trzech obszarów depresji sródsudeckiej (okolice Szczepanowa, Lubawki i Okrzeszyna, rejon Wałbrzycha oraz rejon Nowej Rudy i Słupca). Z rejonu Nowej Rudy i Słupca oraz z rejonów Wałbrzycha i Lubawki znane są flory z utworów wizenu (wczesny karbon), wczesnego namuru oraz westfalu A i B (późny karbon). Poza tym w depresji sródsudeckiej flory karbońskie występują w utworach turneju (wczesny karbon; rejon Wałbrzycha), późnego namuru (rejon Wałbrzycha) i stefanu (późny karbon; okolice Okrzeszyna). Syntetyczne opracowania na temat flor karbońskich Zagłębia podają: Kotasowa i Migier (1983, 1995) oraz Kuchciński (1964). Na nich też oparto poniższe zestawienie.
Według niepublikowanych zestawień dokonanych przez Migier i Dąbrowskiego, na które powołuje się Kotasowa i Migier (1995), makroflora karbońska depresji sródsudeckiej obejmująca przedstawicieli wszystkich głównych grup systematycznych roślin karbońskich liczy około 250 gatunków (widłaki - około 50 gatunków, skrzypy - około 50 gatunków, paprocie właściwe - około 40 gatunków, paprocie nasienne - około 100 gatunków, kordaity oraz przedstawiciele szpilkowych - kilka gatunków).
Z utworów wizenu znanych jest około 70 gatunków, w tym charakterystyczne dla nich widłaki takie jak Sublepidodendron mirabile, Lepidodendron acuminatum, skrzypy takie jak Asterophyllites elegans (ulistnienie kalamita), Sphenophyllum spinulosum czy Sphenophyllopsis culmiana, paprocie właściwe z rodzajów Anisopteris i Rhacopteris oraz paprocie nasienne z rodzajów Acicornopteris, Cardiopteris czy Spathulopteris). Flora wizeńska zawiera szereg gatunków właściwych tylko jej. Część gatunków przechodzi do karbonu późnego, z czego prawie wszystkie kończą swój zasięg w utworach wczesnego namuru (np. Asterocalamites scrobiculatus, Lepidodendron veltheimi, Corynepteris quercifolia czy Neuropteris antecedens). Flora wizeńska została opisana także z Gór Bardzkich (Górecka 1958), a więc z obszaru geograficznie dość bliskiego rejonowi Nowej Rudy.
Flora utworów dolnego namuru Zagłębia liczy około 60 gatunków, w tym charakterystyczne dla niej widłaki z rodzaju Eleutherophyllum, np. E. waldenburgense (epitet gatunkowy pochodzi od niemieckiej nazwy Wałbrzycha - Waldenburg), skrzypy takie jak Calamariophyllum namuriana, szereg gatunków z rodzaju Mesocalamites czy klinolisty z kłosami zarodniowymi klasyfikowanymi w organowym gatunku Sphenophyllostachys waldenburgensis, paprocie właściwe Rhodea goepperti czy Pecopteris aspera, paprocie nasienne takie jak Cardiopteridium waldenburgense, czy szereg gatunków z rodzaju Sphenopteris (np. S. adiantoides ).
Flora utworów westfalu A Zagłębia zawiera gatunki przechodzące z namuru i tu kończące swój zasięg, np. Lepidodendron aculeatum, Wodendron majus (widłaki), Karinopteris acuta, Paripteris gigantea (paprocie nasienne). Liczne gatunki obecne są tylko w utworach westfalu A i B, np. Lepidodendron ophiurus, Sigillaria rugosa, S. scutellata (widłaki), Calamitina goepperti (skrzypy), Corynepteris coralloides, Renaultia schatzlarensis, Crossotheca schatlzarensis (paprocie właściwe), czy liczne paprocie nasienne, np. Sphenopteris nummularia, Mariopteris glabra. Inne gatunki charakterystyczne są tylko dla utworów westfalu A, np. Sphenophyllum amplum (skrzypy) czy Boweria schatzlarensis (paprocie właściwe). Z utworów westfalu A znany jest gatunek Ginkgophyton delvali będący przedstawicielem klasy Ginkgopsida (miłorzębowych). Właściwymi tylko dla flory utworów westfalu B Zagłębia są gatunki takie, jak Sphenophyllum majus i S. emarginatum (skrzypy), Sphenopteris striata, czy Lonchopteris silesiaca (paprocie nasienne). W utworach westfalu A i B występują przedstawiciele kordaitów (np. Cordaites principalis, C. borassifolius).
Z utworów stefanu depresji śródsudeckiej znane są zmineralizowane okazy drewna (Dadoxylon schrollianum). Skamieniałe drewno występuje też w utworach westfalu w okolicach Nowej Rudy (Dziedzic 1958).
Poza gatunkami o stosunkowo krótkim zasięgu stratygraficznym spotykamy w profilu utworów karbonu Zagłębia gatunki bardziej długowieczne, jak np. Lepidodendron lycopodioides (wizen - westfal B), Lepidodendron vortheni (wczesny namur - westfal B), czy organowe taksony widłaków takie jak Lepidostrobophyllum lanceolatum i Stigmaria ficoides (wizen - westfal B). Niewykluczone, że długowieczne gatunki organowe, takie jak wymienione wyżej pnie lepidodendronow czy stigmarie (np. S. ficoides, która w utworach karbonu Górnośląskiego Zagłębia Węglowego występuje od wizenu po westfal D), w rzeczywistości reprezentują szereg osobnych biologicznych gatunków widłaków.
Badania paleobotaniczne Zagłębia Dolnośląskiego rozpoczęły się już w XIX wieku, były wtedy prowadzone przez paleobotaników niemieckich. Z utworów karbonu opisano wówczas szereg nowych gatunków; niektóre z nich wymieniono w powyższym zestawieniu. Większe zbiory flory karbońskiej z Zagłębia znajdują się w kilku instytucjach, między innymi w Muzeum w Wałbrzychu. W oparciu o te ostatnie wydane zostały dwa atlasy (Sagan 1981, 1983). Osady wieku karbońskiego z Zagłębia były też przedmiotem licznych opracowań palinologicznych, mających zastosowanie stratygraficzne (syntetyczne zestawienie znajduje się np. w tomie Prac PIG pod redakcją Zdanowskiego i Żakowej (1995).
Literatura
Bojkowski, K. & Porzycki, J. (eds) 1983. Geological Problems of Coal Basins in Poland. 0-000, Instytut Geologiczny, Warszawa.
Bossowski, A., Ihnatowicz, A., Mastalerz, K., Kurowski, L., & Nowak G.J. 1995. Lithostratigraphy and sedimentologic-paleogeographic development. Sudetes and Fore-Sudetic area. Intra-Sudetic Depression. In: Zdanowski, A. & Żakowa, H. (eds) The Carboniferous System in Poland. Prace PIG 148, 142-147.
Dziedzic, K. 1958. O nowym stanowisku skrzemieniałych pni w karbonie górnym niecki sródsudeckiej. Rocznik Polskiego Towarzystwa Geologicznego 28, 427-434.
Gabzdyl, W. 1994. Geologia złóż węgla. Złoża świata. 0-000. Polska Agencja Ekologiczna, Warszawa.
Górecka, T. 1958. Flora dolnokarbońska Gór Bardzkich. Biuletyn IG 129, 159-197.
Kotasowa, A. & Miger, T. 1983. The role of macroflora in the biostratigraphy of Carboniferous rocks in Polish coal basins.. In: Bojkowski, K. & Porzycki, J. (eds) Geological Problems of Coal Basins in Poland. 130-164. Instytut Geologiczny. Warszawa.
Kotasowa, A. & Migier, T. 1995. Macroflora. In: Zdanowski, A. & Żakowa, H. (eds) The Carboniferous System in Poland. Prace PIG 148, 56-65.
Kuchciński, J. 1964. Podstawowe zagadnienia stratygrafii makroflorystycznej utworów karbońskich niecki śródsudeckiej. Przegląd geologiczny 12, 345-347.
Sagan, E. 1981. Atlas roślin karbońskich na podstawie zbiorów Muzeum Okręgowego w Wałbrzychu. Rośliny widłakowe (Lycophyta). 0-000. Muzeum Okręgowe w Wałbrzychu, Wałbrzych.
Sagan, E. 1983. Atlas roślin karbońskich na podstawie zbiorów Muzeum Okręgowego w Wałbrzychu. Rośliny skrzypowe (Equisetophyta). 0-000. Muzeum Okręgowe w Wałbrzychu, Wałbrzych.
Zdanowski, A. & Żakowa, H. (eds) 1995. The Carboniferous System in Poland. Prace PIG 148, 0-000
Roślinność karbonu na tle flor paleozoiku
GRZEGORZ PACYNA
Uniwersytet Jagielloński
Karbon był okresem panowania paprotników. Był to jedyny okres w lądowej historii roślin, kiedy paprotniki zdominowały większą część krajobrazu. W rozległych zwrotnikowych terenach wilgotnych i bagnach paprotniki świetnie się rozwijały. Najbardziej znane rośliny karbońskie: lepidodendrony, sigilarie i kalamity nie były drzewami, lecz ogromnymi bylinami, o zielonych pniach, utworzonych głównie z żywych tkanek kory wtórnej. Igłowate lub płasko-równowąskie liście drzewiastych widłaków wyrastały z charakterystycznych, spiralnie lub równolegle ułożonych wzgórków, utworzonych przez zewnętrzne partie kory, zwanych poduszeczkami liściowymi. W stanie kopalnym najczęściej spotyka się fragmenty kory ze spłaszczonymi poduszeczkami liściowymi. Różnice w ich morfologii stanowią podstawę odróżniania poszczególnych taksonów. Poduszeczka liściowa paleozoicznych widłaków była swoistym dla nich organem, nie mającym żadnego odpowiednika u dzisiejszych widłaków. Jej funkcją było zaopatrywanie tych roślin w wodę, a także przewietrzanie wnętrza ogromnego pnia. Dolny biegun pnia rozgałęział się kilkakrotnie dychotomicznie, tworząc bardzo charakterystyczny twór zwany stigmarią. Były to grube, prawie poziomo biegnące jakby konary. Miały budowę pędową, nie były więc korzeniami, ale organami korzeniotwórczymi, tzw. ryzoforami. Z nich wyrastały korzenie przybyszowe - appendices, pełniące rolę korzeni oddechowych. Odpadając pozostawiały charakterystyczne owalne blizny. W stanie kopalnym najczęściej są spłaszczone, czasami zachowują się jako trójwymiarowe odlewy, mogą mieć przyczepione appendices.
Puszcze, które porastały wówczas obszar przyrównikowy, w którym znajdowała się Europa i Ameryka Płn., dały początek ogromnym złożom węgla. Okresowo zalewane równiny porastały jednogatunkowe zbiorowiska drzewiastych widłaków z rodzaju Lepidophloios, które osiągały wysokość do 40 m. Tworząc koronę rozwidlały się one wielokrotnie na coraz drobniejsze gałęzie, pokryte drobnymi liśćmi, wyrastającymi z poduszeczek liściowych, których szerokość przewyższała długość. Lepidophloios miał wyszukane organy rozrodcze. Pojedyncza duża makrospora była otoczona przez fałdy tkankowe liścia zarodnionośnego. Całość funkcjonowała jak zalążki roślin nasiennych. Po opadnięciu z roślin macierzystych, takie "zalążki", opatrzone specjalnym "żagielkiem" były roznoszone przez wodę. Żyjące w nieco suchszych miejscach lepidodendrony z rodzajów Anabathra, Flemingites, Lepidodendron i Diaphorodendron były mniejszych rozmiarów. Tworzyły organy rozrodcze w postaci kłosów złożonych z liści opatrzonych makro- i mikrosporangiami. Ich poduszeczki liściowe były dłuższe niż szersze.
Na wyniesieniach rozwijały się bujne zespoły roślinne o większym, niż równiny zalewowe, taksonomicznym zróżnicowaniu, zdominowane przez drzewiaste sigilarie, paprocie nasienne i paprocie właściwe. Sigilarie różniły się od lepidodendronów słabszym rozgałęzieniem korony, a ich poduszeczki liściowe były ułożone na pniu w pionowych, równoległych rzędach. Paprocie nasienne były roślinami nagozalążkowymi, pod względem zróżnicowania taksonomicznego były dominującą grupą w lasach karbońskich. Ich liście, zachowujące się najczęściej jako uwęgliny pozbawione organów rozrodczych, są jednak nieodróżnialne od liści paproci właściwych.
Obrzeża zbiorników porastały zarośla roślin skrzypowych, głównie kalamitów. Różne ich rodzaje zajmowały odmienne siedliska, np. Stylocalamites był formą pionierską, zapoczątkowującą sukcesję roślinną na podłożu utworzonym ze świeżo zdeponowanych osadów i rósł w kępowych skupieniach raczej poza typowymi torfowiskami. Kalamity osiągały kilkanaście metrów wysokości, miały pędy podzielone na węzły i międzywęźla, intensywnie przyrastały na grubość. Ich liście miały blaszki niepodzielone, są opisywane pod nazwami Annularia i Asterophyllites. Przodkowie kalamitów - archeokalamity, które dominowały głównie we wczesnym karbonie, miały podzielone liście. Zielnymi formami skrzypowych były klinolisty o liściach klinowatych, zebranych w okółki.
Zupełnie inne, słabo zresztą poznane zespoły roślinne cechowały obszary wyżynne. Tam dominowały rośliny nagonasienne z grupy kordaitów. Były to wysokie (do 30 m) drzewa o wysmukłych, dochodzących do jednego metra średnicy pniach, niosących na szczycie rozgałęzioną koronę. Młode gałęzie korony pokryte były równowąskimi liśćmi, dochodzącymi do metra długości i 20 cm szerokości. Niektóre ich gatunki tworzyły jednak pionierskie zarośla mangrowiowe. Na wyżynach występowały również rośliny iglaste, o pokroju współczesnej araukarii, które są zaliczane do rodzaju Walchia i Ernestiodendron.
Zmiany klimatu z końcem karbonu położyły ostateczny kres zdominowanym przez paprotniki lasom, nastąpiło wymieranie większości taksonów. W Europie we wczesnym permie resztki tej flory utrzymały się w wilgotniejszych miejscach. W Polsce taka flora znana jest z Karniowic koło Krakowa. Koniec karbonu i początek permu były okresem dużego zróżnicowania florystycznego. W okolicach równika rosły wówczas lasy podobne, jak we wcześniejszych epokach karbonu. Północne obszary zajmowały lasy z dominacją kordaitów, jednak złożone z innych gatunków niż te, które występowały w Eurameryce. Na półkuli południowej (Gondwanie) rozwijała się flora glossopterisowa. Późny perm to zapowiedź nowej ery - mezofitycznej, w której wczesnych okresach dominowały rośliny nagonasienne, a szczególnie szpilkowe.
Ilustrowany tekst wystąpienia zamieszczony został w przewodniku Kursu.
Flora mezozoiku jako tło dla roślin kopalnych z Krasiejowa
ANNA MARIA OCIEPA
Uniwersytet Jagielloński
W mezozoiku w porównaniu z paleozoikiem skończyło się panowanie paprotników i paproci nasiennych. Nie było już drzewiastych widłaków - sigilarii i lepidodendronów; nie było olbrzymich skrzypów - kalamitów; mniejszy był udział paproci. Rośliny widłakowe miały stosunkowo niewielu przedstawicieli. Były to już przeważnie rośliny zielne. Na uwagę zasługują triasowy rodzaj Pleuromeia (2 m wysokości) i kredowy Nathorstiana, które swą budową łączyły cechy sigilarii i dziś występujących poryblinów. Rosnące także u nas w oligotroficznych jeziorach porybliny są więc w prostej linii potomkami sigilarii. Ogólnie w mezozoiku panowała jedność flory, ale zaznaczają się jeszcze pewne różnice między lądem południowym Gondwaną, a północnym Laurazją.
Ilustrowany tekst wystąpienia zamieszczony został w przewodniku Kursu.
Flora kajpru
DANUTA ZDEBSKA
Uniwersytet Jagielloński
Trias (kajper - najmłodsza jego epoka) rozpoczął mezofityczną erę dziejów roślinności na Ziemi. Nastąpiła (zapoczątkowana już w permie) zmiana klimatu. Stał się ciepły i suchy wyniszczając higrofilną roślinność paleozoiku i dając miejsce dla ekspansji, lepiej do nowych warunków przystosowanych, grupom roślin nagozalążkowych.
W dolnym triasie reprezentowane są jeszcze rośliny nawiązujące do karbońskich olbrzymów. Są to rośliny widłakowe z rodzaju Pleuromeia i skrzypowe z rodzaju Schizoneura i Neocalamites dorastające do 2 m wysokości. Z węzłów tych roślin wyrastały jeszcze podługowate liście. W kajprze występują już formy zielne podobne pokrojem do dzisiejszych skrzypów ze zredukowanymi liśćmi. Najbardziej charakterystycznym rodzajem był rodzaj Equisetites, a z widłaków rodzaj Lycopodites.
Nieliczni są też przedstawiciele paproci nasiennych, które wymarły całkowicie w jurze. Charakterystycznym gatunkiem dla kajpru jest Lepidopteris ottonis. Znajdowane są liście podwójnie pierzaste oraz organy rozmnażania żeńskiego - zalążki umieszczone na tarczkach skupionych na rozgałęzionych osiach tworzących rodzaj "kwiatów".
W epoce tej dużą rolę odgrywały paprocie zarodnikowe liczne w rodzaje i gatunki z rodzin reprezentowanych współcześnie we florach tropikalnych. Rodzina Marattiaceae z najliczniejszym rodzajem Danaeopsis o dużych, do 1 m długich, pierzastych liściach. Rodzina Matoniaceae z rodzajem Phlebopteris o liściach tworzących w zarysie wachlarz. Rodzina Dipteridaceae z rodzajem Dictyophyllum o lirowatych liściach z siateczkowatą nerwacją rzadko występującą u paproci.
Najbardziej jednak charakterystycznymi przedstawicielami były rośliny nagozalążkowe z grupy sagowców, miłorzębowych i szpilkowych. Sagowce przypominały współczesne formy rodzaju Cycas. Najbardziej znaną skamieniałością jest Leptocycas gracilis o pniu do 1,5 m wysokim, z pióropuszem liści na szczycie.
Ważną grupą obok sagowców i szpilkowych, współtworzącą charakterystyczną dla tej epoki florę, są pod względem liczby gatunków miłorzębowe. Znajdowane liście o różnym pokroju blaszki należą głównie do rodzaju Ginkgoites.
Najlepiej do zmienionego klimatu przystosowały się rośliny iglaste. Obok panujących w dolnym i środkowym triasie przedstawicieli rodziny Voltziaceae z rodzaju Voltzia na znaczeniu zyskują przedstawiciele wielu nowych rodzin mających współcześnie reprezentantów we florach tropikalnych. Są to przedstawiciele rodzin Araucariaceae (najczęściej spotyka się fragmenty zsylifikowanego drewna rodzaju Araucarites), Cupressaceae z rodzajami Brachyphyllum czy Pagiophyllum i Podocarpaceae z rodzajem Rissikia. W skałach zachowują się w postaci ftagmentów ulistnionych gałązek, w postaci szyszek lub pojedynczych łusek nasiennych.
W środkowym kajprze pojawia się również nieznana dotychczas grupa roślin nagozalążkowych - benetytów, których organy rozmnażania zebrane są w obupłciowy "kwiat". Pokrojem i liśćmi przypominały sagowce. Grupa ta staje się panującą aż do kredy, później wymiera. W kajprze najczęściej znajdowane są ich pierzaste liście określane nazwą rodzajową Ptilophyllum i Anomozamites.
Flora, która wykształciła się w tej epoce, trwała z małymi zmianami w jej składzie aż do kredy. Panującymi roślinami były nagozalążkowe. Trias to również okres, w którym na lądzie Gondwana wymiera flora glossopterisowa a na jej miejsce wkracza flora pochodząca z kontynentów półkuli północnej. Był to okres, w którym zatarła się granica, poprzedno wyraźna, między roślinnością kontynentów półkuli północnej a kontynentem Gondwany półkuli południowej.
Ilustrowany tekst wystąpienia zamieszczony został w przewodniku Kursu.
Małże kajpru Krasiejowa
MICHAŁ BRODACKI
Uniwersytet Warszawski
Pospolite w późnym triasie środkowej Europy małże oznaczane jako Unionites nie zostały dotąd szczegółowo opisane. W dolnej warstwie kościonośnej w Krasiejowie występują one masowo w postaci ośródek i odcisków, towarzysząc szczątkom metopozaurów i fitozaurów. W górnej warstwie są bardzo rzadkie, tam jednak zachowane są wapienne muszle.
Dotychczas zebrane okazy nie przekraczają 3 cm długości, występują w dwóch postaciach: (1) wydłużonej i eliptycznej oraz (2) krótkiej i jajowatej. Analiza morfologii i anatomii nie wyklucza hipotezy, że są to rzeczywiście formy z rodziny Unionidae. W szczególności układ i proporcje odcisków przyczepów mięśni widocznych na ośródkach oraz słabo zachowane zarysy zębów zawiasowych nie różnią się od obserwowanych u współczesnego rodzaju Unio.
Znaczenie okresu triasowego w ewolucji ryb
ŁUKASZ PAŚKO
Uniwersytet Wrocławski
Najstarszym rekinem jest znany tylko z pojedynczych zębów Leonodus z najwcześniejszego dewonu Hiszpanii. Prawdopodobne łuski ryb spodoustych znane są z późnego syluru, a może nawet późnego ordowiku. Cechy pierwotnych spodoustych to m.in. prawie terminalny otwór gębowy i palatoquadratum mocno związane z neurocranium. Wczesne spodouste były aktywnymi drapieżnikami niewielkich rozmiarów, pierwotnie pelagicznymi, odżywiającymi się rybami i głowonogami. Szybko zajęły jednak szereg bentonicznych habitatów przechodząc na odżywianie opancerzonymi bezkręgowcami i kręgowcami. Pierwotnie morskie, szybko opanowały również słodkowodne środowiska.
Powstanie nowoczesnych rekinów (Neoselachii) łączyło się ze zwiększeniem kinetyki czaszki, poprzez oswobodzenie palatoquadratum i przekształcenie zawieszenia łuku żuchwowego, przesunięcie otworu gębowy na spód głowy, uporządkowanie przebiegu włókien w przynajmniej dwuwarstwowym enameloidzie i zwapnienie trzonów kręgów. Zwiększyło to sprawność aparatu szczękowego i zwiększyło możliwości pływackie. Najstarsze takie rekiny (Palaeospinax) pochodzą z końca środkowego triasu. Zadecydowały to o wykorzystaniu przez Neoselachii nowego źródła pokarmu w postaci ryb Holostei. Pierwsze Neoselachii były przybrzeżnymi morskimi drapieżnikami, lecz w kredzie przeszły również do polowania na otwartych wodach. W późnej jurze powstały rekiny o płaskich tnących zębach z piłkowanymi krawędziami. Podobną konstrukcję (zapewne efekt konwergencji) miały zęby ze środkowego dewonu Antarktydy (Mcmurdodus).
Najstarsi przedstawiciele bazalnych promieniopłetwych (Chondrostei) pochodzą z późnego syluru Chin i Skandynawii, ale kompletne szkielety dopiero ze środkowego dewonu Szkocji i Kanady i późnego dewonu Australii. Charakteryzuje je obecność drobnych, romboidalnych łusek ganoidowych, często połączonych stawowo między sobą, płetwa ogonowa epicerkalna i pojedyncza płetwa grzbietowa. Pierwotne promieniopłetwe były niewielkimi morskimi i słodkowodnymi drapieżnikami, o mało funkcjonalnym w wodzie nożycowym aparacie szczękowym (wytwarzającym niewielki efekt zasysania pokarmu przy otwieraniu pyska) i niewielkiej sile zamykania szczęk. W karbonie i permie, częściowo triasie, miało miejsce różnicowanie grupy paleonisków, nieliczne doczekały wczesnej jury; dziś są reliktowe (Polypterus). Nowościami ewolucyjnymi były u nich prawie symetryczna płetwa ogonowa, której górny płat pozbawiony jest w zasadzie struny grzbietowej, pionowa orientacja kości gnykowożuchwowej oraz palatoquadratum, umożliwiające zwiększanie objętości jamy gębowoskrzelowej i zwiększenie masy przywodziciela żuchwy, oraz obecność wyrostka dziobastego na żuchwie, usprawniającego otwieranie szczęk. Cechy te zwiększyły funkcjonalność aparatu szczękowego i lokomotorycznego. Jedna z linii dała początek rybom Holostei.
Pierwsze Holostei znane są z późnego permu Anglii. Ich modyfikacje adaptacyjne obejmują usprawnienie aparatu szczękowego przez rozbicie szkieletu policzka na osobne mobilne kości, umożliwiające znaczne zwiększanie rozmiarów jamy gębowoskrzelowej, wytworzenie stawu między maxillare a podniebieniem, umożliwiajacego rotację tylnej części maxillare ku przodowi, i powstanie nowego mechanizmu opuszczania żuchwy, dzięki połączeniu wyrostka dziobastego z wierzchołkiem preoperculare. W dalszych etapach zwiększająca się ruchliwość palatoquadratum wymusiła utratę jego połączenia z podstawą czaszki. Większość progresywnych cech występuje u bazalnych Holostei w różnych kombinacjach sugerując złożoną historię ewolucyjną grupy. Wczesne Holostei były bardziej ruchliwe i sprawniej pływające od paleonisków, co odbyło się kosztem redukcji środków obrony pasywnej. Zasiedlały one różne środowiska wód morskich i słodkich. Początki różnicowania bazalnych Holostei sięgają triasu a jego szczytowa faza przypadła na jurę. Z triasu znane są trzy rodziny. Wśród nich semiodontidy z późnotriasowych i wczesnojurajskich słodkowodnych jezior grupy Newark (wschodnie wybrzeże Ameryki Północnej) dostarczają dowodów intensywnej specjacji, porównywalnej do niedawnej radiacji pielęgnicowatych z jezior Afryki. Największa grupa monofiletyczna bazalnych nowopłetwych znana z całego mezozoiku, obejmuje m.in. Parasemionotidae, Caturidae, Amiidae i Ophiopsidae. Spośród Parasemionotidae lub bliskiej im grupy wywodzi się prawdopodobnie przodek najbardziej zróżnicowanej gatunkowo grupy kręgowców - ryb kościstych (Teleostei).
Najstarsze szczątki Teleostei pochodzą z późnego triasu Austrii. Ewolucja wczesnych Teleostei objawiała się w dalszej modyfikacji aparatu szczękowego i ogona. Wśród wielu innych cech obejmuje to rozszerzenie myodomu na basioccipitale, dwie kości nadszczękowe, uralne łuki nerwowe zmodyfikowane jako tzw. uroneuralia, mobilne uzębione premaxillare, quadratojugale połączone z quadratum i zewnętrznie symetryczną płetwę ogonową. Najwcześniejsze znane z materiału kopalnego Teleostei były małymi, smukłymi rybami o drapieżnym trybie życia. Główne ich różnicowanie miało miejsce dopiero we wczesnej kredzie a następnie na granicy kredy i trzeciorzędu.
Bliska przodka mięśniopłetwych (Sarcopterygii) może być Psarolepis z przełomu syluru i dewonu Chin. Mięśniopłetwe były pospolitym składnikiem faun morskich i słodkowodnych późnego paleozoiku i mezozoiku. Wczesne Sarcopterygii charakteryzują płetwy parzyste i niektóre nieparzyste osadzone na umięśnionych trzonkach, zaopatrzonych w śródszkieletowe elementy wspierające pochodzące z radialiów, dwie płetwy grzbietowe, łuski kosminowe i neurocranium zaopatrzona w umożliwiający jego wewnętrzną ruchomość staw śródczaszkowy. Pierwotnie były morskimi drapieżnikami, zasiedlającymi strefę przydenną. Później zasiedlały również płytkie, często niedotlenione wody słodkie.
W słodkowodnym triasie występowały trzonopłetwe Mawsoniidae (Actinistia) i dwudyszne z rodzin Gnathorhizidae, Ceratodontidae i Neoceratodontidae. Actinistia charakteryzują się specyficznym typem kinezy czaszki, utratą warstwy kosminowej łusek, redukcją kości szczękowych i przedszczękowych. Dla dwudysznych typowe jest redukcja skórnych kości policzka, zrost palatoquadratum z neurocranium, wytworzenie dwuseryjnego śródszkieletu płetw parzystych, obecność częstych w stanie kopalnym wachlarzowatych płytek zębowych. Większość linii ewolucyjnych Dipnoi i Actinistia wygasła przed końcem kredy.
Kapitosaury
DOROTA MAJER
Uniwersytet Opolski
Kapitosaury należą do grupy płazów tarczogłowych, zwanych również labiryntodontami. Były to duże, drapieżne płazy o długości 3 metrów lub więcej. Ciało ich miało masywną budowę. Głowa ich była duża i szeroka. Występowały od późnego palezoiku do końca triasu. W trakcie ewolucji tyłu czaszki kapitosaurów następowało stopniowe zamykanie się wcięcia usznego. Najbardziej zaawansowane ewolucyjnie pod tym względem są cyklotosaury (Cyclotosaurus). Wcięcie uszne jest u nich całkowicie zamknięte przez połączenie kości łuskowej (o. squamosum) z tabulare (o. tabulare). Mogła być w nim rozpięta błona bębenkowa.
Kapitozaury zamieszkiwały prawdopodobnie brzegi zbiorników wodnych. Trybem życia były zbliżone do dzisiejszych aligatorów. O ich lądowym trybie życia może świadczyć zanik kanału linii bocznej, która jest narządem czucia skórnego charakterystycznym dla zwierząt żyjących w środowisku wodnym. Ponadto przeniesienie oczu bliżej szczytu czaszki zapewniało większe pole widzenia oraz mogło chronić oczy przed uszkodzeniami.
Najdawniej opisanym kapitosaurem, rozpoznanym na podstawie podniebienia i przodu czaszki, był Capitosaurus arenaceus Münster, 1836. W Polsce szczątki kapitosaurów są znane ze śląska Opolskiego z morskich osadów wapienia muszlowego (Kunisch 1890) i z ladynu Tatr, gdzie znaleziono kapitosaura Tatrasuchus kulczyckii (Kotański 1996; Maryańska & Shishkin 1996).
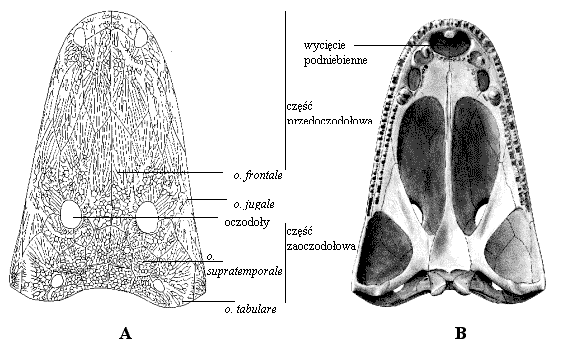 | | Ryc. 1. Rekonstrukcja czaszki kapitosaura Cyclotosaurus sp. znalezionego w Krasiejowie; dach (A) i spód (B) czaszki (rekonstrukcja wykonana przez Tomasza Suleja). |
W Krasiejowie znaleziono dotąd jedynie lużne kości czaszki niezidentyfikowanego gatunku kapitosaura. Jest to bez wątpienia cyklotosaur, na co wskazują całkowicie zamknięte wcięcia uszne przekształcone w otwór owalny. Jednak brak całej nieuszkodzonej czaszki powoduje, że określenie gatunku na tym etapie badań jest niemożliwe. Poniżej przedstawiono porównanie podstawowych cech czaszek cyklotosaurów opisanych dotychczas z terenu Europy. Oprócz czaszki znaleziono również charakterystyczne kręgi zachowujące się w stanie kopalnym jako klinowate w profilu, półokrągłe elementy trzonów (rachitomowe) oraz kość skublową (cleithrum).
Niewątpliwie kapitosaury były istotnym elementem fauny krasiejowskiej i spodziewać się można większej ilości znalezisk, które pozwolą na identyfikację rodzajową i gatunkową.
Literatura
Kunisch, H. 1890. Labirynthodontentereste des oberschlesischen Muschelkalks. Zeitschrift der Deutche geologische Gesellschaft 42, 377-385.
Maryańska, T. & Shishkin, M.A. 1996. New cyclotosaurid (Amphibia: Temnospondyli) from the Middle Triassic of Poland and some problems of interrelationships of capitosauroids. Prace Muzeum. Ziemi 43, 53-83
Schoch, R.R. 2000. Biogeography of stereospondyl amphibians. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 215, 201-231
Welles, S.P. & Cosgriff, J. 1965. A revision of labirynthodont family Capitosauroidae and a description of Parotosaurus peabodyi n. sp. from the Wupatki Member of the Moenkopi Formation of northern Arizona. 1-150. University of California Press, Berkeley and Los Angeles.
Metopozaury
TOMASZ SULEJ
Instytut Paleobiologii PAN
Metopozaury są grupą płazów tarczogłowych znaną od połowy XIX wieku z osadów kajpru niemieckiego. Od tamtego czasu znaleziono liczne nagromadzenia ich kości w Ameryce Płn., Azji i Afryce. Wszędzie występują w osadach późnego triasu w stosunkowo wąskim przedziale czasu - w późnym karniku i wczesnym noryku. W wiekszosci stanowisk występują z fitozaurami i aetozaurami. Razem z jeszcze innymi już mniej licznymi kręgowcami tworzyły stosunkowo kosmopolityczny zespół dużych kręgowców.
Kolejnym miejscem występowania tego zespołu w Europie jest niedawno odkryte stanowisko w Krasiejowie na śląsku Opolskim (Dzik et al. 2000). Można wyróżnić w nim dwa główne poziomy kościonośne, różniące się udziałem poszczególnych grup płazów i gadów. W poziomie górnym dominują zwierzęta lądowe: aetozaury, cyklotozaury oraz dwunożne roślinożerne archozaury. W poziomie dolnym natomiast przeważają wodne metopozaury i fitozaury. Tylko w tym poziomie spotyka się też szczątki ryb ganoidowych i dwudysznych.
Metopozaury były płazami osiągającymi prawie 3 m długości. Miały dużą płaską czaszkę stanowiącą ok. 1/4 długości ciała. W widoku z góry miała ona typowy dla płazów tarczogłowych paraboliczny kształt. Stosunkowo wyjątkowe było natomiast położenie oczodołów, które znajdowały się w przedzie czaszki tuż przy jej brzegu (u większości płazów oczodoły położone byly z tyłu czaszki, blisko siebie). Czaszka metopozaurów była urzeźbiona na całej górnej powierzchni. Silnie wyodrębnione z rzeźby byly kanały linii bocznej, w których za życia zwierzęcia występowały komórki zmysłowe wrażliwe na ruch opływającej zwierze wody. Tak wyraźne kanały wskazują na to, że nie były pozbawionymi funkcji rudymentami. Dowodzą zatem wodnego trybu życia metopozaurów (Milner 1994).
świadczą o tym także nieskostniałe elementy śródręcza i śródstopia, które pozostawały chrzęstne przez całe życie zwierzęcia i nie zachowały się w znanych z Maroka całych szkieletach metopozaurów (Dutuit 1976). Prawdopodobnie żyjąc w środowisku wodnym płazy te nie obciążały tych części kończyn i dlatego nie uległy one skostnieniu. Jest to zjawisko dosyć często spotykane wśród płazów tarczogłowych (Milner 1994).
U wszystkich przedstawicieli tej grupy na piersi występowały duże płyty kostne: dwa obojczyki i międzyobojczyk. U metopozaurów kości te były stosunkowo grube i ciężkie. Masywność tych kości nie mogła być przystosowaniem do przenoszenia dużych obciążeń przy chodzeniu, bo pozostaje to w sprzeczności ze słabym szkieletem kończyn. Mogły więc pełnić funkcje obciążników pomagających utrzymać się pod wodą zwierzęciu oddychającemu przecież powietrzem atmosferycznym. Pas biodrowy metopozaurów był w dużej mierze chrzęstny. Częściowo kostniały tylko kości biodrowa i kulszowa. Panewka stawowa była w przewazajacej czesci chrzęstna. Przy takiej budowie miednicy można wykluczyć lądowy tryb życia tych zwierząt.
Wobec tylu argumentów wskazujących na życie w srodowisku wodnym metopozaurów zagadkowe jest całkowite skostnienie trzonów kręgów (intercentrum), które poza ogonem są cylindryczne. Jest to stosunkowo rzadkie wśród plazów tarczoglowych, nawet w grupach uważanych za bardziej lądowe (np. u występującego w Krasiejowie cyklotozaura). Niewielkie i rzadko znajdowane u metopozaurów pleurocentra prawdopodobnie najczęściej pozostawały chrzęstne (Dutuit 1976).
Metopozaury stanowią najliczniej reprezentowaną w Krasiejowie grupę zwierząt. Posiadamy 12 całych czaszek, liczba kłykci potylicznych w mniej kompletnych okazach dokumentuje ok. 50 osobników. Równie liczne ich występowanie jest notowane w stanowiskach z Ameryki Północnej (Colbert & Imbrie 1956).
Stanowiska metopozaurów najbliższe Krasiejowa znajdują się w Wirtembergii, niedaleko Stuttgartu. Rezultaty porównania pochodzących stamtąd okazów z krasiejowskimi są przygotowywane do publikacji. Kluczowa ideą jest nowa interpretacja położenia lacrimale względem oczodołów u opisanego z Wirtembergii Metoposaurus diagnosticus. Cecha ta jest uważana za jedną z najważniejszych w systematyce metopozaurów. Do tej pory sądzono, że gatunek ten miał lacrimale oddzieloną od krawędzi oczodołów przez szew jugale i praefrontale (Fraas 1889). Jednakże badanie holotypu, jak i nie opisanych okazów znalezionych później, nie potwierdziło tego poglądu. Wydaje się bowiem, że podobnie jak u większości metopozaurów, lacrimale była podłużna. Jej kształt i kontakt z krawędzią oczodołów w holotypie jest taki sam jak u okazów pochodzących z Krasiejowa. Także kształt i rzeźba claviculi i interclaviculi są bardzo podobne. Na tej podstawie uznano, że metopozaury występujące w Krasiejowie należą do gatunku Metoposaurus diagnosticus. Dokładne pomiary parietale wykazały pewne różnice między populacją z Wirtembergii i Krasiejowa. Różnią się one wartością kąta zawartego między szwem łączącym lewe parietale i supratemporale, a szwem łączącym ich prawe odpowiedniki (kat parietale). Nie wydaje się jednak, by była to różnica pozwalająca wyodrębnić te populacje jako podgatunki. Nowe dane dotyczece jedynego znanego z Europy metopozaura wskazuja na koniecznosc odrzucenia cechy dotyczacej polozenia lacrimale jako odrózniajacej ten rodzaj od form znanych z Ameryki Pólnocnej (Hunt 1993). Jedyna cecha rózniaca rodzaj Metoposaurus od amerykanskeigo Buettneria pozostaje rzezba interclaviculi (Colbert & Imbrie 1956).
Wszystkie znane z Europy metopozaury mają taki sam kształt urzeźbienia interclaviculi. Jest to płaska szeroka tarcza pokryta od strony brzusznej licznymi dołkami i rowkami. Dołki znajdują się w jej centralnej części, skad stopniowo wydłużają się w promieniscie rozchodzące się rowki. U Metoposaurus diagnosticus pole pokryte dołkami zajmuje stosunkowo niewielką powierzchnię. U gatunków pochodzących z Ameryki Północnej proporcje te są natomiast odwrócone - w urzeźbieniu interclavicul dominują dołki. Wydaje sie, ze pierwotniejszy stan tej cechy wystepuje u Metoposaurus diagnosticus, który jest najstarszym znanym metopozaurem. Wystepuje już w formacji Schilfsandstein, głęboko poniżej Blasensandstein, gdzie z metopozaurami wciąż występują prymitywne fitozaury Paleorhinus. Poza Europą gatunki z rodziny Metoposauridae występuja także w Indiach w formacji Malerii (Chowdhury 1965), w Maroku w formacji Timesgadiouine (Dutuit 1976), w licznych formacjach Texasu, Nowego Meksyku, Pensylwanii i Wyoming (Long & Murry 1995) oraz formacji Isalo II na Madagaskarze (Dutuit 1978).
Tak szerokie rozprzestrzenienie metopozaurów wyróżnia je spośrod innych płazów tarczogłowych póznego triasu takich, jak cyklotozaury, mastodonzaury, brachiopoidy, plagiozaury oraz almazaury. Czesc z nich wspólwystepowala w pojedynczych stanowiskach z metopozaurami: trematozaury Mastodonsaurus i Hyperokynodon i cyklotozaury w Wirtembergii (Fraas 1889), kapitosauroida byc moze również na Madagaskarze (fragment podniebienia; Lucas 1998); latiskopidy Almasaurus w Maroku (Dutuit 1976).
Powstaje zatem pytanie, czemu tylko metopozaury są jedynymi kosmopolitycznymi płazami tarczogłowymi w póznokarnijsko-wczesnonoryckim zespole dużych kręgowców, do którego należą także aetozaury i fitozaury. Wydaje się, że nałożyły się na to dwa czynniki:
(1) Przede wszystkim niezwykłe rozprzestrzenienie pewnego typu biotopu w dużych śródlądowych zbiornikach wodnych powstalych w wyniku podniesienia poziomu morza w późnym karniku (Aigner & Bachman 1998), w warunkach panujacego wtedy ciepłego i wilgotny klimatu z cyklami powodzi i suszy (Simms et al. 1995; Kirby 1993). Podstawową adaptacją metopozaurów do życia w takim środowisku własnie duze rozmiary i oczodoły umieszczone w przodzie płaskiej, parabolicznej, stosunkowo dlugiej czaszki, blisko jej krawędzi, były. Płaskie zwierzę polujące z zasadzki leżąc przy dnie posiadając dobrze rozwiniete kanały linii bocznej i oczodoły umieszczone z przodu czaszki może szybciej zlokalizowac potencjalna ofiarę (Sulej 1998).
(2) To, że metopozaury wykształciły swoiste przystosowania we właściwym czasie - przed rozprzestrzenieniem się właściwego im środowiska życia. Najstarsze metopozaury znane są już z regresywnych osadów Schilfsandstein środkowego karniku. Ponieważ zadne inne późnotriasowe płazy tarczogłowe nie miały takich przystosowań, one wlasnie wygrały konkurencję. ściślej lądowe cyklotozaury być może również występowały kosmopolitycznie tworząc z aetozaurami zespół kręgowcow ściśle lądowych, a nie znajdujemy ich szczątków, ponieważ jako zwierzeta lądowe miały mniejsze szanse na fosylizację. Aetozaury sa liczniej reprezentowane w materiale kopalnym jako zwierzeta roślinożerne, o wyższej produkcji biologicznej, niż drapieżne cyklotozaury.
Literatura
Aigner, T. & Bachman, G. H. 1998. Sequence stratigraphy of the Germanic Triassic: A short overview. Hallesches Jahrbuch für geowissenschaften Reihe B 6, 23-26.
Chowdhury, T. R. 1965. A new metoposauroid amphibian from the Upper Triassic Maleri Formation of central India. Philosophical Transactions of the Royal Society of London B 250, 1-52.
Colbert, E. H. & Imbrie, J. 1956. Triassic Metoposaurid amphibians. Bulletin of the American Museum of Natural History 110, 403-452.
Bolt, J. R. & Chatterjee, S. 2000. A new temnospondyl amphibian from the Late Triassic of Texas. - Journal of Paleontology 74, 670-683.
Dzik, J., Sulej, T., Kaim, A. & Niedźwiedzki, R. 2000. Późnotriasowe cmentarzysko kręgowców lądowych w Krasiejowie na śląsku Opolskim. Przegląd geologiczny 48, 226-235.
Dutuit, J. M. 1976. Introduction a l'etude paleontologique du Trias continental marocain. Description des premiers Stegocephales recueillis dans le couloird'Argana (Atlas occidental). Memoires du Museum National d'Histoire Naturelle, Paris, Serie C 36, 1-253.
Dutuit, J. M. 1978. Descripion de quelques fragments osseux provenant de la région de Folakara (Trias supérier malagache). Bulletin de Museum Nationale d'Histoire Naturelle, Series III 516, 79-89.
Fraas, E. 1889. Die Labyrinthodonten aus der Schwäbischen Trias. Palaeontographica 36,1-158.
Hunt, A. P. 1993. Revision of the Metoposauridae (Amphibia: Temnospondyli) and description of a new genus from western North America. Museum of Northern Arizona Bulletin 59, 67-97.
Ingavat, R & Janvier, P. 1981. Cyclotosaurus cf. posthumus Fraas (Capitosauridae, Stereospondyli) from the Huai Hin Lat Formation (Upper Triassic), northeastern Thailand. Geobios 14, 711-725.
Lucas, S. G. 1998. Global Triassic tetrapod biostratigraphy and biochronology. Paleogeography, Paleoclimatology, Paleoecology 143, 347-384.
Kirby, R. E. 1993 Relatonship of Late Triassic basin evolution and faunal replacement events in the southwestern United States: Perspectives from the upper part of the Chinle Formation in Northern Arizona. New Mexico Museum of Natural History & Science Bulletin 3, 233-242.
Milner, A.R. 1994. Late Triassic and Jurassic ampibians: fossil record and phylogeny. In: N.C. Fraser & H.-D. Sues (eds) In the Shadow of Dinosaurs. 5-22. Cambridge University Press.
Simms, M.J., Ruffell, A.H., & Johnson, A.I.A. 1995. Biotic and climatic changes in the Carnian (Triassic) of Europe and adjacent areas. In: N.C. Fraser & H.-D. Sues (eds) In the Shadow of Dinosaurs. 352-365. Cambridge University Press.
Sulej, T. 1998. Geneza typu przystosowawczego wielkich jeziornych czworonogów póznego triasu. Praca magisterska, Wydział Biologii Uniwersytetu Warszawskiego.
Watson, D. M. S. 1958. A new labyrinthodont (Paracyclotosaurus) from the Upper Trias of New South Wales. Bulletin of the British Museum (Natural History) 3: 233 - 263.
Fitozaury
MARIUSZ LUBKA
Uniwersytet Wrocławski
Fitozaury były mieszkańcami późnotriasowych rozlewisk, jezior i rzek. Wszędzie gdzie występowały - a rozprzestrzenienie ich było niemal kosmopolityczne - występowały licznie. Były dużymi zwierzętami osiągającymi do 12 metrów długości, co stawia je w rzędzie największych drapieżników swoich czasów. Te ziemnowodne gady uznaje się powszechnie za ekologicznych odpowiedników dzisiejszych krokodyli.
Przystosowania anatomiczne. - Fitozaury porównywano do krokodyli od czasu odkrycia w pierwszej połowie XIX wieku. Istotnie, fitozaury i nowoczesne krokodyle mają prawie identyczny plan budowy: wydłużoną głowę, nieco spłaszczone ciało zaopatrzone w krótkie, acz mocne kończyny, długi silny ogon. Te podobieństwa są jednak powierzchowne i mają z pewnością naturę konwergencji, nie zaś bliskiego pokrewieństwa. Budowa kości oraz ślady przyczepów mięśni sugerują, iż kończyny fitozaurów były prymitywniejsze niż u krokodyli. Za życia były zapewne podciągnięte pod tułów i ustawione kolumnowo. Zachowane tropy fitozaurów wskazują, że zwierzęta te mogły poruszać się unosząc ciało wysoko nad ziemią oraz że nie wlokły za sobą ogona. Charakteryzowały się obecnością pancerza złożonego z umiejscowionych wzdłuż kręgosłupa parzystych językowatych tarczek, drobnych tarczek okrywających podgardle, licznych tarczek rozrzuconych w skórze kończyn oraz gęsto ułożonych żeber brzusznych. Nie jest pewne, czy ogon fitozaurów pokryty był pancerzem.
Najbardziej oczywiste różnice, w porównaniu do krokodyli, występują w obrębie czaszki. Można tu wymienić przesunięcie zewnętrznych otworów nosowych fitozaurów na szczyt czaszki pomiędzy oczy, podobnie jak ma to miejsce u waleni czy zauropodów. Związana z tą cechą jest budowa rostrum. U krokodyli przednia część pyska składa się z kości szczękowych (nozdrza znajdują się na końcu pyska), u fitozaurów wydłużeniu podlegały kości przedszczękowe "spychając" nozdrza ku tyłowi. Choć u fitozaurów zaznacza się rozwój wtórnego podniebienia, nigdy nie uzyskało ono takiej formy, jak u nowoczesnych krokodyli. Zapewne jednak posiadały podniebienie miękkie umożliwiające im oddychanie i połykanie zdobyczy w tym samym czasie, lub pod wodą.
U fitozaurów można wyróżnić trzy różne formy wykształcenia czaszki: (1) charakteryzujący się długim i wąskim pyskiem (np. Mystriosuchus), (2) u zwierząt o pyskach stosunkowo krótkich i wysokich (np. Smilosuchus) oraz (3) stanowiący formę pośrednią między poprzednimi (np. Paleorhinus). Podobne zróżnicowanie można obserwować u nowoczesnych krokodyli. W parze z budową czaszki szedł u fitozaurów typ uzębienia. Z długim pyskiem wiąże się uzębienie jednorodne. Taki wydłużony pysk, stosunkowo słabe - i nie licząc lepiej rozwiniętych zębów na samym końcu pyska - niezróżnicowane uzębienie wskazują na rybożerność. Rolą zębów było przede wszystkim przytrzymywanie ofiary oraz odpowiednie ułożenie jej przed połknięciem, nie zaś rozszarpywanie jej. Do tego celu nadawało się z kolei zróżnicowane uzębienie fitozaurów o wysokich, spłaszczonych z boku pyskach. W ich masywnych szczękach umożliwiających wytworzenie sił zdolnych zabić dużą ofiarę tkwiły zęby, z których przednie, potężnie rozwinięte zadawały śmiertelne rany, leżące za nimi umożliwiały przetransportowanie pożywienia ku tyłowi pyska, gdzie znajdowały się z kolei zęby pozwalające na rozdarcie ciała ofiary na możliwe do połknięcia kawałki. Zachowana zawartość żołądków okazów z rodzajów Paleorhinus i Pseudopalatus wskazuje, iż istotnie fitozaury żywiły się niewielkimi czworonogami, z których część to formy typowo lądowe, a rozpoznanie tafonomiczne fauny ze Stubensandstein w Niemczech pokazało, że duże fitozaury były w stanie poradzić sobie z ofiarą większą niż one same.
Ekologia i biogeografia. - Szczątki fitozaurów występują często w osadach słodkowodnych, co jednoznacznie wskazuje, iż zwierzęta te związane były ze stałymi zbiornikami wodnymi: jeziorami czy rzekami. Nieliczne fragmenty szkieletów znalezione w osadach morskich uznawane były zazwyczaj za allochtoniczne. Niedawne znaleziska dowiodły, że późne, noryckie fitozaury występowały nie tylko w przybrzeżnych wodach lagunowych, ale że również zapuszczały się na otwarte morze.
Pierwsze znane człowiekowi okazy fitozaurów pochodzą z Niemiec. Do dziś ich szczątki znaleziono też w innych krajach europejskich: we Francji, Szwajcarii, Austrii, Włoszech, a ostatnio i w Polsce. Bogatego materiału dostarczyły odkrycia dokonane we wschodniej i południowo-zachodniej części Stanów Zjednoczonych. Ponadto fitozaury znane są z Maroka, z Tajlandii oraz z Indii, a dyskusyjne szczątki również z Madagaskaru. Słabo zachowany, budzący wątpliwości co do swego fitozaurowego pochodzenia (nawiasem mówiąc najprawdopodobniej zaginiony) materiał pochodzi z Chin. Jednym słowem fitozaury miały szeroki zasięg występowania, nawet jeżeli weźmiemy pod uwagę, że omawiane stanowiska nie były tak bardzo od siebie oddalone ok. 215 Ma temu, jak dziś. Karnijski rodzaj Paleorhinus został znaleziony na obszarze części Stanów Zjednoczonych, w południowych Niemczech, Austrii, Polsce, Maroko i w Indiach, miał zatem rozprzestrzenienie niemal kosmopolityczne. Należy tu zaznaczyć, iż żaden z równowiekowych czy późniejszych rodzajów nie zasiedlał już tak rozległego obszaru. W noryku zespoły faunistyczne, w których skład weszły fitozaury miały węższe zasięgi. I tak na przykład rodzaj Mystriosuchus czy Nicrosaurus znany jest tylko z Europy, podczas gdy w tym samym czasie występował w Ameryce Północnej autochtoniczny rodzaj Pseudopalatus. Brak jednak fitozaurów w niektórych stanowiskach, w których szczątki późnotriasowych faun lądowych występują w obfitości. I tak, nie znaleziono szczątków fitozaurów w Południowej Afryce ani w Ameryce Południowej. Powody takiego stanu rzeczy nie są jasne; mają zapewne związek z panującym tam bardziej suchym i chłodniejszym klimatem. Ogólnie rzecz ujmując wszystkie stanowiska, w których znaleziono fitozaury lokują się między 30oN i 30oS (przy układzie lądów u schyłku triasu).
Historia ewolucyjna. - Rząd Phytosauria reprezentuje (wraz z aetozaurami, rauizuchami oraz krokodylami i ich krewnymi) krokodylową gałąź archozaurów (Crurotarsii). W dawniejszej nomenklaturze fitozaury zaliczano do dużej i bardzo zróżnicowanej, obecnie nieformalnej, grupy tekodontów. O ile stosunek Phytosauria do innych grup archozaurów został stosunkowo dobrze poznany i nie budzi wątpliwości, o tyle relacje wewnątrz rzędu nadal pozostają niejasne. Nie osiągnięto jak dotąd konsensusu w kwestii liczby rodzajów i rodzin. Wynika to z dwóch podstawowych przyczyn.
Po pierwsze, fitozaury były grupą szybko ewoluującą. Trudno więc o niezależną i dostatecznie precyzyjną korelację wiekową ich znalezisk. Trudno określić, które populacje, mniej lub bardziej odległe w czasie czy przestrzeni, należą do jednego gatunku. Nader często kluczowe cech taksonomiczne bywają zaciemniane przez zmienność. Zmienność owa może być odzwierciedleniem dymorfizmu płciowego, zmian wzrostowych, albo po prostu indywidualnych cech osobniczych, patologii spowodowanych zranieniami, czy w końcu stanu zachowania skamieniałości.
Po drugie, rozległe rozprzestrzenienie geograficzne przedstawicieli grupy utrudnia badania i porównania materiałów muzealnych. Najczęściej łączy się wszystkie znane formy w rodzinę Phytosauridae, czasem z wyłączeniem Paleorhinus jako pierwszej grupy zewnętrznej (w podejściu kladystycznym). Ponieważ nie można dostrzec w omawianej sprawie choćby cienia porozumienia (i, jak się wydaje, długo go jeszcze nie będzie) nie będę rozważać szczegółów klasyfikacji. Nie sposób też ustalić liczby rodzajów; zapewne jest ich około ośmiu. Szczątki fitozaurów krasiejowskich należą do stosunkowo pierwotnego rodzaju Paleorhinus i być może reprezentują dobrze znany ze stanowisk północnoamerykańskich gatunek P. bransoni.
Pochodzenie fitozaurów nie jest znane. Najstarsze, karnijskie ich szczątki należą do rodzaju Paleorhinus, formy o wyraźnie już ukształtowanych cechach rzędu. Należy się zatem spodziewać, że grupa ma swoje korzenie co najmniej w środkowym triasie. Jest kilka doniesień o szczątkach fitozaurów z okresu wcześniejszego niż późny trias (pstry piaskowiec i wapień muszlowy), jest to jednak materiał niezwykle skąpy i - jeśli rzeczywiście należy do jakichś form związanych z fitozaurami, co budzi wiele kontrowersji - nie mówi praktycznie niczego o wczesnej ich ewolucji. Jedynie czaszka, pochodząca jakoby z pstrego piaskowca (wczesny trias) Bernburga (Niemcy) opisana przez Otto Jaekela w 1910 r. jako Mesorhinus fraasi, była na tyle kompletna, że mogłaby dostarczyć informacji na temat pochodzenia i ewolucji fitozaurów, jednak została zniszczona podczas II Wojny światowej. Pozostał tylko szczegółowy opis. Jedni autorzy traktują M. fraasi jako ważny gatunek i budują na tej podstawie teorie na temat ewolucji fitozaurów, według innych poważne wątpliwości budzi nie tyle pokrewieństwo okazu bernburskiego z fitozaurami, co datowanie skamieniałości, nawiasem mówiąc ustalone przez Jaekela w drodze złożonej dedukcji. Najlepszym rozwiązaniem wydaje się zaniechanie teoretyzowania na podstawie omawianego okazu i cierpliwe oczekiwanie na nowe znaleziska dokumentujące pochodzenie i wczesną ewolucję fitozaurów. Korzeni grupy można by dopatrywać się wśród Proterochampsidae, rodziny prymitywnych gondwańskich "tekodontów", u których zaznacza się przesunięcie zewnętrznych otworów nosowych ku tyłowi, tak charakterystyczne dla fitozaurów. Najstarsi przedstawiciele tej grupy znani są ze ladynu Argentyny. Nie wiemy, kiedy dokładnie pojawiły się wczesne fitozaury. Najprawdopodobniej było to w późnym karniku (rodzaj Paleorhinus), za czym przemawiałby fragment czaszki znaleziony w dobrze datowanych morskich osadach triasu alpejskiego.
Fragmenty szkieletów należące niewątpliwie do fitozaurów znane są tylko z górnego triasu. Większość autorów jest zdania, że Phytosauria to grupa, która powstała u zarania późnego triasu, szybko się rozwijała zyskując nawet status skamieniałości przewodnich, i z końcem tego okresu wymarła bezpotomnie. Koniec karniku, noryk oraz retyk to czas, kiedy fitozaury, szybko ewoluując, bujnie się rozwijały. Przywiązane do jednego - ziemnowodnego - środowiska wykształciły szereg wyspecjalizowanych form. Stały się częstym składnikiem części ówczesnych faun odgrywając w nich niebagatelną rolę, a wszędzie gdzie się pojawiały występowały w dużej liczbie osobników. Na początku noryku paleorinusy zostały zastąpione przez liczne nowe i bardziej wyspecjalizowane rodzaje. U schyłku triasu zakończyła się jednak historia grupy. Dobrze prosperujące w opanowanych przez siebie środowiskach, a co więcej zdobywające środowiska nowe (morskie), fitozaury znikły nagle z zapisu kopalnego. Dwa przypadki odkrycia szczątków fitozaurów w dolnej jurze (Chiny - słabo zachowane szczątki czaszki, zaginione; wiek kontrowersyjny; Francja - trzy zęby w hettangu, brak pewności, czy szczątki autochtoniczne) to zbyt skąpe dowody na to, iż fitozaury przeżyły koniec triasu w weszły do jury.
Aetozaury
TOMASZ SULEJ
Instytut Paleobiologii PAN
Aetozaury są grupą archozaurów występującą w lądowych osadach karniku i noryku na wszystkich kontynentach oprócz Australii i Antarktydy. Były to czworonożne zwierzeta roślinożerne, pokryte pancerzem skladajacym sie z prostokatnych tarczek, które są najczęściej występującymi szczątkami tych zwierząt. Formy prymitywne osiagaly 3 m dlugości, znane są jednak gatunki małe, u których dorosłe osobniki miały zaledwie 1 m (Aetosaurus) jak i bardzo duże np. Desmatosuchus, który osiagał 6 m dlugości. Pancerz składał się z czterech rzędów tarczek na grzbiecie i 6-8 po stronie brzusznej. Tarczki podobnie jak u wszystkich archozaurów były urzeźbione i miały na środku górnej powierzchni wyniosłość, z której u niektórych gatunków powstały duże kolce. Ponieważ tarczki nachodziły na siebie dachówkowato, przedni brzeg wykształcony był w postaci listwy. Przyśrodkowe tarczki grzbietowe były szerokie i miały ostre wierzchołki. Tarczki boczne natomiast miały mniej regularne i zazwyczaj zaokrąglone brzegi. To właśnie kształt i forma urzeźbienia tarczek są diagnostycznymi cechami w omawianej grupie. Jest tak nie tylko dlatego, że istotnie sa charakterystyczne dla poszczególnych gatunków, ale także dlatego, że pozostałe elementy szkieletu są znajdowane bardzo rzadko.
Do tej pory jedyna w calosci zachowana czaszka aetozaura nalezy do Aetosaurus ferratus z Wirtembergii. Dobrze zachowanej czaszki nie udało się znalesc także podczas prac poszukiwawczych w Krasiejowie pod Opolem (Dzik et al. 2000), gdzie występuja one w dwóch poziomach kościonośnych. Nagromadzenie kości co najmniej czterech osobników (dowodzą tego cztery prawe kości dentale) znalezione zostało w poziomie górnym, w którym dominują szczątki zwierząt lądowych. W poziomie dolnym natomiast tarczki aetozaurów są stosunkowo nieliczne, bowiem dominuja tam zwierzęta wodne - metopozaury i fitozaury. Udało się jednak znalezć fragment lewej i prawej kości dentale, należącej prawdopodobnie do jednego osobnika.
Do tej pory wypreparowano tylko część przywiezionego z Krasiejowa materiału, w tym wiele tarczek. Tarczki przyśrodkowe przykrywające szyję sa gładkie lub mają bardzo delikatną rzeźbę. Wyniosłość jest zaznaczona jako niewielka wypukłość. Rzezba wszystkich tarczek grzbietowych staje się wyraźniejsza wraz ze zbliżaniem się do okolicy krzyżowej, gdzie ma postać wyodrębnionych, promieniście rozchodzących się rowków i dolków. Także wyniosłość w tej części jest wykształcona jako równoległy do osi ciała ostry grzbiet. Przednia listwa jest wyraźna we wszystkich tarczkach, poza brzusznymi, których przednia krawędź jest tylko lekko zawinieta. Poniewaz podobne tarczki występują u Stagonolepis robertsoni z Lossiemouth Sandstones ze Szkocji (Walker 1961), materialy z Krasiejowa zaliczono do tego rodzaju, znanego również z Teksasu, Nowego Meksyku, Arizony (Long & Murry 1995), Argentyny (Heckert & Lucas 1999) i Wirtembergii (Hunt & Lucas 1992). Walker (1961) przedstawil rekonstrukcje czaszki oraz calego zwierzęcia wykonane na podstawie kilku fragmentów czaszki i szkieletu pozaczaszkowego i analogii z Aetosaurus ferratus z Wirtembergii.
Znalezione w Krasiejowie dwa dobrze zachowane fragmenty tylnej części czaszki oraz całe kości premaxillare, maxillare, nasale i jugale umozliwiły zrekonstruowanie czaszki. Czaszka z Krasiejowa jest wieksza i bardziej masywna. Rózni sie tez liczba zębów w żuchwie - w Krasiejowie 7 zębów, 9-10 u S. robertsoni. Nie zaznaczone na rekonstrukcji Walkera wystepy na parietale (widoczne z góry) znane są u wszystkich aetozaurów; w opisywanym przez niego materiale prawdopodobnie nie zachowały sie.
Mimo, ze budowa czaszki wielu gatunków aetozaurów pozostaje nieznana, cechy istotne do odtwarzania trybu zycia tych zwierzat sa dobrze udokumentowane. Bezzębny przód zuchwy pokryty był prawdopodobnie rogowym dziobem, który w połączeniu z bezzebnym rozszerzonym ryjkiem sugeruje przystosowanie do rycia w ziemi. Potwierdzaja to duże krętarze na kościach przedramienia, które u zwierzat grzebiacych zwiekszaly sile konczyn pomocnych przy tym sposobie poszukiwania pokarmu (Parrish 1994). Nie mógł to być jednak pokarm twardy, bowiem stożkowate zęby aetozaurów, mające bardzo slabo rozwinięte piłkowanie, nie noszą wyraźnych sladów ścierania. Aetozaury były prawdopodobnie zwierzętami roślinożernymi jedzącymi miekkie liście i wygrzebującymi z ziemi kłącza.
Aetozaury pojawiły się w osadach karniku już jako formy posiadające wszystkie właściwe sobie cechy. Choć nie ma jeszcze zgodności co do ewolucji aetozaurów wydaje się, że Stagonolepis jest najprymitywniejsza znana forma. Jego tarczki byly urzezbione radialnie podobnie jak innych archozaurów i nie mialy wyraznych kolców. Lossiemouth Sanstones ze Szkocji, w których wystepuje Stagonolepis robersoni jest zapewne najstarszym stanowiskiem zawierajacym aetozaury. Podstawą takiego datowania jest współwystepowanie rynchozaura Hyperokynodon wystepującego takze w formacji Maleri z Indii, z której pochodza metopozaury i fitozaury z rodzaju Paleorhinus, wystepujace takze w Krasiejowie. Rynchozaury ze Szkocji i Indii róznia sie od siebie tylko jedna malo znaczaca cecha: proporcja miedzy szerokoscia bocznego i srodkowego pola zebowego na maxilli (Benton 1990). H. gordoni ze Szkocji ma wezsze boczne pole zebowe, podobnie jak starsze rynchozaury, natomiast u H. huxleyi z formacji Maleri proporcja ta jest odwrócona, co moze wskazywac na jego większe zaawansowanie ewolucyjne. Sugeruje to, ze osady Lossiemouth Sanstones sa starsze, niz formacja Maleri i równowiekowy z nią Krasiejów.
W Ameryce Pólnocnej i Wirtembergii Stagonolepis współwystępuje z fitozaurami z rodzaju Rutiodon, który jest bardziej zaawansowany niz Paleorhinus. Z nadleglych warstw Wirtembergii pochodzi sławne nagromadzenie calych szkieletów malych aetozaurów Aetosaurus ferratus. Rodzaj ten znany jest takze z noryckich osadów Alp Wloskich, z Nowego Meksyku, New Jersey i Pólnocnej Karoliny w Ameryce Pólnocnej oraz z Grenlandi (Lucas et al. 1998). Choc Walker (1961) ze wzgledu na niewielkie rozmiary i ksztalt zebów uznal je za najprymitywniesze aetozaury, wydaje się jednak, ze sa one potomkami Stagonolepis. Prawdopodobnie najmlodszym przedstawicielem tej linii jest Neoaetosauroides z formacji Los Colorados z Argentyny (Lucas et al. 1998). Jeszcze w karniku wyksztalcila sie z niej duza grupa bardziej zaawansowanych aetozaurów.
Gałąź ewolucyjną równolegla do linii Stagonolepis-Neoaetosauroides tworzyła grupa, której przodkowie wytworzyli kolce na silnie wygietych tarczkach bocznych i utracili zeby na premaxilli (Parrish 1994). Najstarszym znanym przedstawicielem tej grupy jest Longosuchus wspólwystepujacy z fitozaurem Paleorhinus w Teksasie i Nowym Meksyku w Ameryce Pólnocnej (Long & Murry 1995). Z mlodszych osadów pochodzi Desmatosuchus, majacy dlugie kolce na tarczkach szyjnych i wpustowe polaczenie tarczek. Bardzo szerokie tarczki z urzezbieniem w postaci dolków wytworzyly blisko z nim spokrewnione Paratypothorax i Typothorax o tulowiu w ksztalcie dysku (Parrish 1994).
Literatura
Benton, M. J. 1990. The species of Rhynchosaurus, a rhynchosaur (Reptilia, Diapsida) from the Middle Triassic of England. Philosophical Transactions of the Royal Society of London B 328, 213-306.
Dzik, J., Sulej, T., Kaim, A. & Niedźwiedzki, R. 2000. Późnotriasowe cmentarzysko kręgowców lądowych w Krasiejowie na śląsku Opolskim. Przegląd geologiczny 48, 226-235.
Heckert, A. B. & Lucas, S. G. 1999. A new aetosaur (Reptilia: Archosauria) from the Upper Triassic of Texas and phylogeny of aetosaurs. Journal of Vertebrate Paleontology 19, 50-68.
Hunt, A. P. & Lucas, S. G. 1992. The first of the aetosaur Parathypothorax andressi (Reptilia: Archosauria) in the western United States and its biochronological significance. Paläontologische Zeitschrift 66, 147-157.
Long, R. A. & Murry, P. A. 1995. Late Triassic (Carnian and Norian) tetrapods from the southwestern United States. New Mexico Museum Natural History Science Bulletin 4, 1-254.
Lucas, S. G., Heckert, A. B. & Huber, P. 1998. Aetosaur (archosauromorpha) from the Upper Triassic of the Newark Supergroup, eastern United States, and its biochronological significance. Palaeontology 41, 1215-1230.
Parish, J. M. 1994. Cranial osteology of Longosuchus meadei and the phylogeny and distribution of the Aetosauria. Journal of Vertebrate Paleontology 14, 196-209.
Walker, A. D. 1961. Triassic reptilies from the Elgin area: Stagonolepis, Dasygnathus and their allies. Philosophical Transactions of the Royal Society of London 244, 103-204.
Dinozaury triasu
JERZY DZIK
Instytut Paleobiologii PAN
Pochodzenie dinozaurów nie jest dostatecznie udokumentowane paleontologicznie. Głównym tego powodem są trudności z korelacją warstw skalnych zawierających najstarsze szczątki dinozaurów i ich fragmentaryczność.
Do niedawna za najstarsze i równocześnie najprymitywniejsze dinozaury uchodziły drapieżne gatunki z rodzajów Eoraptor i Herrerasaurus z formacji Ischigualasto w Argentynie, z którymi współwystępuje bardzo fragmentarycznie poznany roślinożerny Pisanosaurus, prawdopodobny przodek dinozaurów ptasiomiednicznych. W obrębie Ameryki Płd. możliwa jest korelacja wiekowa późnego triasu w oparciu o stowarzyszone z dinozaurami rynchozaury i stąd wiadomo, że geologicznie nieco starszy jest drapiezny dinozaur Staurikosaurus z górnej części formacji Santa Maria w Brazylii. Wydaje się jednak, że te południowoamerykańskie dinozaury nie dokumentują rzeczywiście pierwszych stadiów ewolucji swojej grupy. Próby powiązania standardowego podziału kontynentalnego późnego triasu północnej Ameryki z resztą świata doprowadziły bowiem do wniosku, że fragmentaryczne szczątki dinozaurów z formacji Salitral w Nowym Meksyku i formacji Popo Agie w Wyoming są wyraźnie starsze. Współwystępują z fitozaurem Paleorhinus, uważanym za skamieniałość przewodnią dla późnego karniku.
Paleorhinus występuje też w Krasiejowie. Wraz z nim znaleziona został przed kilkoma laty fragmentaryczna kość krzyżowa dwunożnego archozaura przypominająca odpowiednią część szkieletu Staurikosaurus. W trakcie ubiegłorocznych wykopalisk w górnym poziomie kościonośnym udało się znaleźć około czterystu kości tego zwierzęcia, reprezentujących stado co najmniej siedmiu osobników, w tym cztery niekompletne szkielety. Pozwoliło to na wykonanie pełnej rekonstrukcji szkieletu. Jego miednica, z w pełni skostniałą panewką, ma cechy znane tylko u bardzo pierwotnych dinozaurów (Guibasaurus) i pierwotniejszych od nich rauisuchów, w szczególności ladyńskiej Bromsgroveia. Budowa kręgów szyjnych jest nieco bardziej zaawansowana, niż u późnoladyńskiego Marasuchus z fauny Los Chanares Argentyny i bliska ceratozaurowi Liliensternus z noryku Niemiec. śląski gad był jednak roślinożerny. Najbliższy mu anatomicznie pod względem budowy czaszki jest prosauropod Thecodontosaurus z noryku Anglii. Szczególną cechą gatunku z Krasiejowa, zbliżającą go do dinozaurów ptasiomiednicznych, jest wydatny ostry dziób na wierzchołku żuchwy.
Okazuje się więc, że zwierzę to wykazuje mozaikowy zestaw cech anatomicznych ustawiający je nie tylko wiekowo, ale i anatomicznie pośrodku pierwszych rozgałęzień drzewa rodowego dinozaurów.
Geneza dwunożności dinozaurów
EWA BARYCKA
Uniwersytet Opolski
Szeroko, prawie poziomo rozstawione kończyny pierwszych płazów ledwie unosiły ciało podczas pełzania po lądzie. Boczne wygięcia ciała pomagały wydłużyć krok. Wyodrębnione w późnym permie gady łuskonośne wciąż wykorzystywały jako siłę napędową mocne przednie kończyny. Boczne wygięcia ciała zwiększały długość kroku, a tylne kończyny redukowały się pełniąc tylko funkcję podporową.
U gadów naczelnych przeciwnie, wzmacniały się tylne kończyny i szybko zwiększyły swoją długość w stosunku do przednich. Powstał złożony, wklęsło-wypukły staw między kością skokową i piętową, przebity otworem na tętnicę, zaś 5 kość śródstopia stała się hakowato wygięta i połączona z 4 tarsale po zaniku 5 tarsale. Dzięki temu gady naczelne mogły podjąć w niedługim czasie próbę wyprostowania postawy czyli podciągnięcia kończyn bliżej ciała. Naczelne z rodziny Erythrosuchidae miały przekształconą panewkę stawu biodrowego tak, że kość udowa nie tkwiła już poziomo lecz pod pewnym kątem w dół, dając w efekcie pół wyprostowaną postawę. Podobną budową charakteryzowała się euparkeria, wczesnotriasowy gad naczelny, który dzięki swoim niewielkim rozmiarom mógł przebiegać niewielkie dystanse tylko na tylnych kończynach, goniąc ofiarę czy uciekając przed niebezpieczeństwem, mimo iż stawy i ułożenie jego kończyn nie były przystosowane do dwunożności. Rozstawione na boki kończyny zwierząt takich jak euparkeria czy Erythrosuchidae wymagały stałego napięcia mięśni i praktycznie uniemożliwiały bieg z wysoko uniesionym tułowiem, gdy przy dwunożności cały ciężar ciała skupiał się na tylnych kończynach. W późnym triasie zmiany w budowie miednicy i stępu umożliwiły całkowicie wyprostowaną postawę, a nawet dwunożność.
W linii ewolucyjnej prowadzącej do krokodyli uformował się skomplikowany staw, w którym kość skokowa ze żłobieniem po otworze na tętnicę tworzyła trzpień pasujący do panewki na kości piętowej z ukośnie przechodzącą płaszczyzną zgięcia między nimi, a powiększony guzek kości piętowej służył jako przyczep ścięgna mięśnia gastrocnemus, zwalniając z tej funkcji hakowatą 5. kość śródstopia. Wciąż najdłuższy czwarty palec wskazywał na niesymetryczne przeniesienie siły, mimo, że kość udowa ustawiona była pionowo, opierając się na grzebieniu górnej część panewki stawu biodrowego na kości biodrowej. Chociaż miały wyprostowaną postawę, właśnie ukośne zgięcie stawu skokowego i filarowe ułożenie kości udowej nie sprzyjało przejściu na dwunożność. Zwierzęta te, głownie związane ze środowiskiem wodnym, powracały do chodu z rozstawionymi kończynami, korzystniejszego w tym środowisku.
W linii małych zwierząt prowadzących do dinozaurów kość skokowa, bez śladu żłobienia, i piętowa bez guzka piętowego zrastały się ze sobą i z dalszymi końcami kości piszczelowej i strzałkowej, przykrytymi od przodu i boku przez szeroki wyrostek kości skokowej, dodatkowo stabilizujący te połączenie. Płaszczyzna zgięcia przechodziła między nimi i kośćmi dolnego piętra stępu, tworząc bardzo symetryczny zawiasowy staw, co uwydatniło się w wydłużeniu trzeciego palca. Wyrostki kości miednicy wydłużyły się tworząc dużą powierzchnię dla przyczepu mięśni. Powstał również masywny grzebień na biodrowej, górnej części panewki stawu biodrowego, a oparta o niego główka była skręcona. Kość udowa wygięta była łukowato do przodu. Kości podudzia śródstopia bardzo się wydłużyły dzięki czemu tylna kończyna stała się dwa razy dłuższa od przedniej. Uformowała się palcochodność, jak u większości szybko biegających zwierząt. Gdy grzebień nad panewką przejął prawie cały ciężar ciała, środkowa część panewki wydelikatniła się i przestała kostnieć, tworząc w skamieniałych szkieletach otwór między kością biodrową i łonową. Otwór ten jest główną cechą diagnostyczną dinozaurów. Wczesne dinozaury były prawdopodobnie fakultatywnie dwunożne, wspierając się podczas postojów na przednich kończynach. Wymieranie późno triasowe stworzyło okazję do rozwoju i wielkiego zróżnicowania, dzięki któremu powstały całkowicie dwunożne jak i czworonożne dinozaury, oraz cały szereg pośrednich sposobów poruszania się, umożliwiając 120 milionów lat panowania dinozaurów na Ziemi.
|