|
Spis treści:
Podstawy nomenklatury taksonomicznej - JERZY DZIK
Problemy nomenklatoryczne na styku taksonomii organowej i biologicznej w paleobotanice - DANUTA ZDEBSKA
Taksonomia i nomenklatura spor i pyłków a systematyka biologiczna - MARIA ZIEMBIŃSKA-TWORZYDŁO
Problemy z doborem typów opisowych w paleontologii kręgowców - MARIUSZ LUBKA
Nomenklatoryczny i taksonomiczny status podgatunku w paleozoologii kręgowców - TOMASZ SULEJ
Nomenklatoryczne konsekwencje typologicznej i populacyjnej koncepcji gatunku w paleozoologii bezkręgowców - MICHAŁ ZŁOTNIK
Dane genetyczne a kryteria morfologiczne gatunku otwornic planktonowych - WOJCIECH MAJEWSKI
Egzekwowanie zasady priorytetu w taksonomii paleozoologicznej - JERZY DZIK
Podstawy nomenklatury taksonomicznej
JERZY DZIK
Instytut Paleobiologii PAN
Po ponad dwu stuleciach powszechnego użycia nie ma już w biologii odwrotu od zasad wprowadzonych przez Carla Linné (standardem jest wydanie jego Systema Naturae z 1758 roku). W dostosowanej do dzisiejszych potrzeb postaci są one podstawą Międzynarodowych Kodeksów Nomenklatury Zoologicznej (ICZN) i Botanicznej (ICBN). Najistotniejszą wspólną cechą nomenklatury taksonomicznej w zoologii i botanice jest uznawanie zasady priorytetu w odniesieniu do nazw, co oznacza, że w użyciu powinna być nazwa jednostki wprowadzona jako pierwsza. By ograniczyć możliwe niejednoznaczności, gatunki opiera się na wzorcowych okazach, które muszą być zdeponowane w publicznych instytucjach muzealnych.
Wzorzec gatunku
Każdy gatunek biologiczny ma wyznaczony wzorcowy okaz (holotyp) o znanym miejscu pochodzenia, które jest stanowiskiem typowym locus typicus (w paleontologii ponadto warstwą typową stratum typicum). Umożliwia to identyfikację, w razie potrzeby, typowej populacji. Wszelkie dane odnoszące się do tej populacji mogą być użyte do wskazania różnic w stosunku do innych gatunków. Treść diagnozy gatunku nie jest w jakimkolwiek stopniu wiążąca i nawet jeśli autor gatunku wskazał zupełnie fałszywie jego rzekome charakterystyczne cechy, o tym czy gatunek jest odrębny czy nie, decydują wyłącznie cechy populacji typowej a nie subiektywna ocena biologów. Stąd nierzadko się zdarza, że nazwy gatunkowe uznane za młodsze synonimy, są przez kolejnych rewidentów przywracane do użycia. Przeważnie holotyp jest jedynie częścią organizmu (zasuszoną gałązką z kwiatostanem, szkieletem bądź skórą) i listy synonimów, szczególnie w paleontologii, mogą zawierać nazwy utworzone w oparciu o zupełnie nieporównywalne szczątki. Może się nawet zdarzyć, że holotyp jest częścią ciała pozbawioną jakichkolwiek cech charakterystycznych. O ile wówczas nie uda się, mimo to, wskazać populacji typowej, nazwa określana jest jako wątpliwa (nomen dubium) i pomijana w dalszych rozważaniach. Dla taksonomów nadrzędną wartością jest jednak stabilność klasyfikacji, jeśli więc jest choćby znaczące prawdopodobieństwo identyfikacji populacji typowej, nawet jeśli holotyp zaginął, nazwę pozostawia się, wskazując w takiej sytuacji zastępcze typy opisowe gatunku. Jest to szczególnie ważne w paleontologii, gdzie dokonuje się rekonstrukcji bardzo niekiedy złożonych szkieletów i lista synonimów może być długa i różnorodna. Podejście do zasad nomenklatury taksonomicznej bywa jednak różne. Paleobotanicy pod osobnymi nazwami opisują kwiaty męskie i żeńskie, liście, odciski kory i drewno kopalnych roślin naczyniowych. Często odchodzą od zasady priorytetu na rzecz oceny, który z organów jest bardziej charakterystyczny. Przeciwstawne podejście charakteryzuje paleozoologów rekonstruujących aparaty gębowe paleozoicznych zwierząt. Tam wartość diagnostyczna poszczególnych elementów aparatu w ogóle nie jest brana pod uwagę przy decyzjach nomenklatorycznych. Decyduje wyłącznie prawdopodobieństwo przynależności poszczególnych okazów do tej samej populacji. Daje to stabilność nomenklatury i bardzo sprzyja postępowi badań ewolucyjnych i paleobiologicznych.
Kodeks nomenklatury zoologicznej nie obdarza uprawnieniami priorytetu wytworów ani śladów działalności organizmów kopalnych. W licznych przypadkach rozróżnienie skamieniałych ciał od wytworów czy skutków aktywności jest jednak trudne do przeprowadzenia. Zdarza się wręcz, że całe wielkie grupy organizmów są rozpoznawane wyłącznie w oparciu o przejawy ich behavioru, bardziej charakterystyczne, niż anatomia (np. lepione z kolagenu kolonie graptolitów, czy sposoby żerowania prymitywnych anatomicznie robaków). Wbrew temu, wśród geologów popularna jest tendencja do traktowania śladów działalności życiowej jako obiektów czysto formalnej klasyfikacji (ichnotaksonomii), nie opartej na metodologii taksonomii biologicznej.
Binomeny linneuszowskie
Każdy gatunek w linneuszowskiej klasyfikacji należy do rodzaju i jego nazwa obejmuje obydwie rangi, rodzaju i gatunku. W nazewnictwie obowiązuje priorytet, ważna jest nazwa (w obrębie każdej z rang z osobna) wprowadzona najwcześniej. Biologowie dla porządku dodają więc do łacińskich nazw rodzaju (z dużej litery) i gatunku (obie wyodrębnione krojem czcionki, w normalnym tekście kursywą) również nazwisko autora gatunku i datę opublikowania pierwszego opisu. Jeśli ktoś później zmieniał przynależność rodzajową gatunku, nazwisko autora z datą umieszcza się w nawiasie, a za nim (w nomenklaturze botanicznej) następuje nazwisko tego, który dokonał uznanego dziś za właściwe przemieszczenia. Należymy zatem do gatunku Homo sapiens Linné, 1758. Nazwa podgatunku (rasy geograficznej czy wycinka linii ewolucyjnej w paleontologii) zawiera jeszcze jeden człon, jest więc trinomenem, np. Homo sapiens neanderthalensis. Zwykle nazwisko i datę rozdziela przecinek (taką postać mają powołania na źródła w niektórych wydawnictwach), ale niektórzy uważają go za zbędny. Jeśli autorów jest dwóch czy trzech podaje się ich nazwiska, jeśli więcej, wystarczy po pierwszym dodać et al. (et alii - łac. i inni). Najprostszym sposobem uniknięcia problemu w jakim języku powinien być łącznik między nazwiskami jest zastosowanie ligatury (podwójnej czcionki) oznaczającej łacińskie et, czyli &.
Pojęcie gatunku biologicznego nie przystaje, z definicji, do organizmów niezdolnych do mejozy. Tradycyjnie określeń binominalnych używa się jednak do wszelkich niskiej rangi klas tych organizmów, nawet bakterii i wirusów.
Taksony ponadrodzajowe
Granice rodzajów, rodzin, rzędów, gromad i typów (w zoologii) oraz klas i gromad (w botanice) są, w przeciwieństwie do granic gatunku, subiektywne. Nie całkiem jednak, bowiem powszechnie się dziś przyjmuje, że wszystkie jednostki zaliczane do taksonu (jednostki systematyki biologicznej) muszą mieć wspólne ewolucyjne pochodzenie, tzn. pochodzą od jednego wyjściowego gatunku należącego do danego taksonu. Jest wiele szkół taksonomii proponujących radykalnie odmienne kryteria jej uprawiania, nie należy to jednak do dziedziny nauki lecz raczej buchalterii biologicznej.
Nazw jednostek wyższej rangi, mimo łacińskiego brzmienia, nie należy wyodrębniać z tekstu kursywą. Naukowa nomenklatura taksonomiczna jest wyłącznie łacińskojęzyczna (z rzadkimi wyjątkami, np. wirusów) i obowiązuje we wszystkich językach. Nazewnictwo w innych językach ma jedynie charakter zwyczajowy. Wprowadzanie równoległych binomenów i zmienianie ich za każdym razem, kiedy taksonomowie zmienią zdanie co do prawidłowej formy łacińskiej nazwy naukowej, pozbawione byłoby przecież sensu. Dlatego zbyteczne jest używanie podwójnych polskich nazw gatunków, jeśli nie ma do tego szczególnej potrzeby.
Zasady tworzenia nazw taksonomicznych
Wszelkie nazwy taksonomiczne muszą być zlatynizowane, przynajmniej przez pozbawienie nieznanych łacinie znaków diakrytycznych. Wymawiać się je powinno w zgodzie z tradycją średniowiecznej łaciny. Nie ma, niestety, żadnych ograniczeń, jeśli idzie o następstwo głosek czy długość nazwy i wiele z nich w gruncie rzeczy niemal nie da się wymówić po łacinie (np. Scheuchzeria czy Yaoxiaognathus).
Nie ma też nieprzekraczalnych reguł tworzenia nazw rodzajowych czy gatunkowych (te ostatnie powinny być w formie przymiotnikowej) a istotne jest jedyne rozpoznanie rdzenia w nazwach o wyraźnie łacińskiej czy greckiej proweniencji. Do rdzenia tego dodaje się bowiem końcówki tworząc nazwy rangi ponadrodzajowej. Opublikowanie takiej nazwy (wraz z diagnozą o dowolnej treści) wskazuje automatycznie rodzaj typowy a tym samym i typowy gatunek (który musi być jednoznacznie wskazany przy wyznaczaniu rodzaju). Oznacza to, że typowym rodzajem rodziny (jak również nad- i podrodziny) Hominidae jest rodzaj Homo (automatycznie) a gatunkiem typowym rodzaju jest Homo sapiens (przez monotypowość, jako jedyny zaliczony do rodzaju przez Linneusza). Jest to jeden z nielicznych gatunków, któremu zaniedbano wyznaczyć holotyp. Zoologiczną końcówką rodzinową jest -idae, botaniczną -aceae. Odpowiednio dla podrodziny jest to -inae (w botanice -idae), dla nadrodziny zoologicznej -oidea (takie jest zalecenie ICZN, ale stale jeszcze spotyka się -acea). Zwykle stosowaną końcówką rzędu w botanice jest -ales, a w zoologii -ida. Wielu zoologów opiera się jednak temu i często stosowaną końcówka rzędu jest -formes (np. w odniesieniu do ryb czy ptaków). Zmiana końcówek nie wymaga uzasadniania merytorycznego i może być dokonywana automatycznie. Często używane w zoologii podrzędy mają końcówki -ina. Do nazw taksonomicznych rangi rzędu nie odnoszą się jednak zasady priorytetu i w przypadku szczególnie dawno i szeroko używanych nazw (Primates, Carnivora, Coleoptera) nie ma potrzeby zmieniania czy też sztucznego opatrywania ich niezgodnymi z duchem nomenklatury końcówkami. Odnosi się to również do botanicznych rodzin (Graminae, Compositae). Stabilność i łatwość posługiwania się nomenklaturą taksonomiczną są ważniejsze od względów doktrynalnych (takson musi być jedynie monofiletyczny w szerokim znaczeniu).
Zasada priorytetu i przypisywania autorstwa odnosi się do wszelkich rang taksonów na poziomie rodzaju, rodziny czy rzędu (tzn. zmiana rangi taksonu z podrodziny na rodzinę, czy nadrodzinę łączy się tylko z automatyczną zmianą końcówki).
Ortografia spolszczeń.- Jest tylko jedna naukowa nomenklatura taksonomiczna i nie ma potrzeby tłumaczenia jej na każdy z języków narodowych z osobna. Jeśli jednak pewne nazwy przechodzą do powszechnego codziennego użycia korzystne może być nadanie im końcówek i ortografii bliższych polszczyźnie. Jest w tym jednak niebezpieczeństwo mimowolnego wprowadzenia się do terminologicznego getta, praktycznie uniemożliwiającego poszerzanie wiedzy przez lekturę literatury obcojęzycznej. Uczenie się podwójnych nazw egzotycznych i kopalnych organizmów byłoby zaś zbytnim, a co więcej zbytecznym, obciążeniem pamięci. Nazwanie w naszym języku wszystkich żyjących współcześnie organizmów (trudna do określenia liczba używanych jednostek rangi ponadgatunkowej wynosi kilkaset tysięcy!) tudzież terminów anatomicznych odnoszących się od organizmów kopalnych (kolejne kilkadziesiąt tysięcy pojęć) nie powinno być ambicją przyrodników.
Przy ubóstwie książek przyrodniczych w języku polskim i publikowaniu niemal wszelkich rezultatów prowadzonych w kraju badań w języku angielskim studiowanie biologii oznacza w istocie lekturę tekstów obcojęzycznych. Dlatego zasadą powinno być wstrzemięźliwie posługiwanie się słowami, które nie mają żadnej korespondencji z terminologią międzynarodową. Jest kilka zasad, którym podporządkować trzeba tworzenie nowych terminów fachowych: (1) na polskie nazwy z pewnością zasługują te wszystkie organizmy, które występują (choćby w cyrku) na szeroko rozumianym terytorium naszego kraju, o ile są dostrzegalne gołym okiem albo mają wpływ na codzienne życie człowieka; (2) nie ma najmniejszej potrzeby wprowadzania takich nazw dla zwierząt egzotycznych lub kopalnych, z którymi stykają się jedynie fachowcy lub entuzjaści; (3) jeśli zachodzi potrzeba nazwania egzotycznego organizmu terminem nienaukowym, roztropniej użyć ludowej nazwy obcojęzycznej używanej w kraju, gdzie występuje, niż tworzonego ad hoc polskiego neologizmu; (4) nie ma powodu, by nie dodawać polskich końcówek do rdzeni nazw łacińskich, jeśli nazwa ma szansę być użyta przez fachowców spoza specjalności, do której przynależy; (5) nie byłoby wówczas roztropne takie przekształcanie ortografii by rdzeń stał się nieidentyfikowalny nawet dla fachowców (np. "fakops" od Phacops lub "perysfynkt" od Perisphinctes) i słuszniej posłużyć się takimi zasadami jak w odniesieniu do obcojęzycznych imion własnych (jak w efemerycznych neologizmach "reaganomika" czy "thatcheryzm", ale również "chopinowski").
Problemy nomenklatoryczne na styku taksonomii organowej i biologicznej w paleobotanice
DANUTA ZDEBSKA
Uniwersytet Jagielloński
Klasyfikacja roślin kopalnych łączy się z licznymi trudnościami i problemami, które są obce botanikom zajmującym się roślinami żyjącymi. Systematyk opisując dzisiejszą roślinę ma ją w całości ze wszystkimi organami, które mają systematyczną wartość. Podstawową trudnością w pracy paleobotanika jest fragmentaryczność znajdowanych kopalnych szczątków roślinnych. Zachowują się w skałach najczęściej w postaci kawałków różnych organów, takich jak liście, fragmenty pędów, drewna, korzenie czy nasiona. Są zmienione pod względem fizycznym i chemicznym. Ulegają bowiem deformacjom, a w pewnych przypadkach po roślinie nie pozostaje nic poza zewnętrznym jej odciskiem na skale.
Ten niekorzystny w wielu przypadkach stan zachowania uniemożliwia dokładne poznanie ich budowy morfologiczno-anatomicznej. Zwykle też fragmenty tych organów nie pozostają ze sobą w łączności morfologicznej, są przemieszane, mogą należeć do różnych rodzajów i gatunków. Ustalenie pierwotnej przynależności szczątków jest więc czynnością trudną i żmudną. Im starsza geologicznie jest dana grupa szczątków kopalnych, tym większe są różnice w stosunku do dziś żyjących roślin i trudności w ustaleniu pozycji systematycznej takich makroszczątków. Rośliny różnią się niejednokrotnie tak, że można je zaliczyć co najwyżej do wyższych jednostek taksonomicznych jak rząd, klasa czy podgromada.
Historycznie rzecz ujmując, paleobotanicy posługiwali się i posługują w pewnym stopniu sztucznym systemem klasyfikacji odnoszącym się do oddzielnych organów występujących w stanie kopalnym. W wyniku tego powstał stosowany do części opisywanych roślin kopalnych specjalny system nomenklatoryczny. Roślinnym szczątkom kopalnym, tak jak roślinom dzisiejszym, nadaje się podwójną nazwę, rodzajową i gatunkową, i grupuje w wyższe jednostki taksonomiczne: rodziny, rzędy, klasy, podgromady. Nazwy te jednak często mają niepełną wartość biologiczną. Nie jest w pełni porównywalna ich diagnoza z diagnozą roślin żyjących. Jest to związane z faktem, że charakterystyka rodzaju rośliny kopalnej często opiera się na jej fragmentach, a niekiedy na jednym tylko organie. W związku z tym w paleobotanice mamy do czynienia najczęściej z tzw. rodzajami formowymi (zwanymi również organowymi lub narządowymi). Tworzone są jednak tak jak rodzaje roślin współczesnych wg przepisów zawartych w Międzynarodowym Kodeksie Nomenklatury Botanicznej (obowiązuje Kodeks z roku 1987, wydanie w języku polskim pochodzi z 1993 r.).
Przykładem rodzajów formowych są: Stigmaria - określa organy korzeniotwórcze widłaków karbońskich, Dadoxylon - drewno rośliny szpilkowej, Lepidophyllum - pojedyncze liście widłaków karbońskich czy Pecopteris - liście paproci z grupy Pteridophylla. W obrębie tych rodzajów opisywane są gatunki, których diagnoza oparta jest na różnicach w budowie morfologicznej czy anatomicznej danego organu, na którego podstawie utworzony został rodzaj formowy.
Większość roślin zbudowana jest z wielu części, powoływanie się więc na całą roślinę wymaga odmiennego systemu nomenklatury. Przy rekonstrukcji całej rośliny, gdy szczęście sprzyja i pozwalają na to znaleziska różnych jej organów, postępuje się wg dwóch metod. Według jednej cały organizm otrzymuje nową nazwę rodzajową; wg drugiej, otrzymuje nazwę organu, który został nazwany pierwszy. Przykładem może być rekonstrukcja pierwszej paproci nasiennej Lyginopteris oldhamia. Nim uzyskano do tego wystarczające podstawy, jej części znane były pod różnymi nazwami rodzajowymi i gatunkowymi o odmiennej przynależności systematycznej: Lyginodendron oldhamia - były to fragmenty łodyg z budową anatomiczną prymitywnych roślin szpilkowych, Sphenopteris hoeninghausi - liście klasyfikowane wśród paproci, Lagenostoma lomaxi - nasiona roślin nagonasiennych, Rachiopteris - ogonki liściowe, Kaloxylon hookeri - korzenie.
Do celów klasyfikacji roślin cechą o dużej wartości systematycznej są organy rozrodcze. Znajdowane wraz z innymi organami są podstawą do utworzenia rodzaju, którego diagnoza jest porównywalna z diagnozą pokrewnego rodzaju dzisiejszego. Roślina taka umieszczana jest w systemie obok dzisiejszych przedstawicieli, jako ich forma wymarła. Klasyfikacja roślin mimo długich studiów i stosowania nowych metod badawczych, daleka jest jeszcze od odzwierciedlania naturalnych pokrewieństw i przebiegu ewolucji. Drogą do poznania historii roślin są badania roślin kopalnych. Najcenniejsze są te rodzaje, których budowa morfologiczno-anatomiczna oraz sposób rozmnażania zostały dobrze poznane. Przykładów takich rodzajów kopalnych porównywalnych z rodzajami współczesnych roślin jest coraz więcej. Utworzenie systemu w pełni naturalnego nie będzie zapewne możliwe. Tylko w nielicznych przypadkach udało się bowiem ustalić związki rodowe większych grup taksonomicznych roślin.
Taksonomia i nomenklatura spor i pyłków a systematyka biologiczna
MARIA ZIEMBIŃSKA-TWORZYDŁO
Uniwersytet Warszawski
Taksonomia i nomenklatura to dwa różne pojęcia, pierwsze oznacza klasyfikację, drugie nazewnictwo, a oba razem stanowią podstawę systematyki, czyli systemu (porządku) roślin. System, który odzwierciedla pokrewieństwa poszczególnych grup roślinnych jest systemem "naturalnym", czyli filogenetycznym.
Ta część palinologii, która zajmuje się sporomorfami (zarodnikami i ziarnami pyłku, czyli organami rozmnażania - pierwszymi komórkami gametofitu), jako odrębna gałąź w obrębie botaniki i paleobotaniki ma istotne znaczenie przy rekonstrukcji filogenezy roślin, a więc dróg, jakimi wędrowały zmieniające się cechy roślin. Pierwszym jednak zadaniem na tej drodze jest poznanie, opisanie i sklasyfikowanie rozproszonych kopalnych sporomorf, z jakimi palinolog ma do czynienia przy opracowywaniu osadów różnego wieku. Czynności tej służy również nadawanie opisanym sporomorfom nazw o zasięgu międzynarodowym, czyli nomenklatura zgodna z aktualnym Międzynarodowym Kodeksem Nomenklatury Botanicznej (ICBN). Potrzeba istnienia ogólnego systemu klasyfikacji jest sprawą oczywistą, ponieważ tylko dzięki uprzedniemu nazwaniu a następnie pogrupowaniu opisywanych sporomorf można je uporządkować i ocenić ich ogromną różnorodność.
Palinolog, opisując sporomorfy występujące w stanie kopalnym, oderwane od rośliny macierzystej, dąży przede wszystkim do stwierdzenia ich przynależności botanicznej do współcześnie żyjącego gatunku, rodzaju lub rodziny. Przy analizie spor i ziaren pyłku z osadów holoceńskich i plejstoceńskich nie ma z tym większych trudności, natomiast już w osadach trzeciorzędowych występują sporomorfy pochodzące z rodzajów wymarłych. Im starsze są osady, tym więcej jest wymarłych wyższych jednostek systematycznych, których spory czy ziarna pyłku przetrwały w stanie kopalnym. Niezmiernie rzadko udaje się powiązać rozproszone sporomorfy z macierzystymi dla nich roślinami kopalnymi.
Dlatego sporomofy kopalne, opisywane z osadów różnego wieku, układane są w systemach opartych wyłącznie na cechach morfologicznych i traktowane jako odrębne części roślin. Określane dla nich gatunki, łączone są w rodzaje. Oczywiście pojęcie gatunku w takim przypadku jest umowne, ponieważ ustanowiony dla rozproszonych sporomorf gatunek może zarówno reprezentować cały rodzaj lub nawet rodzinę roślin wymarłych, jak i nie obejmować nawet jednego gatunku wymarłej rośliny. Podobnie utworzenie wyższej jednostki systematycznej - rodzaju oparte jest na cechach morfologicznych sporomorf. Takie morfologiczne taksony wskazywane były jako "taksony formowe" (form species, form-genus).
Tak więc jedynie ścisły opis morfologiczny stał się podstawą klasyfikacji sporomorf i utworzenia systemów morfologicznych (tzw. systematyki sztucznej), w których do tej samej jednostki systematycznej zalicza się organy roślin nie spokrewnionych ze sobą. Dopiero w następnym etapie poznania sporomorf można próbować odszukania powiązań danego gatunku morfologicznego z rośliną macierzystą. Taka droga jest podstawą wszystkich prac paleobotanicznych.
Skrajne różne poglądy na systematykę sporomorf wyrażało wielu palinologów, głównie radzieckich (np. Zauer, Ananowa i inni), którzy dla rozproszonych sporomorf trzeciorzędowych i mezozoicznych nie tworzyli oddzielnych rodzajów, tylko zaliczali kreowane przez siebie gatunki kopalne do rodzajów dzisiejszych roślin.
Zdaniem wielu palinologów dla celów praktycznych, jak biostratygrafia czy paralelizacja osadów, wystarczająca jest klasyfikacja morfologiczna. Pierwsze systemy morfologiczne zostały wprowadzone przy klasyfikacji megaspor karbońskich (Reinish 1881; Bennie & Kidston 1886). System zaproponowany przez Ibrahima (1933), również ograniczony do karbońskich sporomorf, łączył mikro- i makrospory. Zostały one podzielone na trzy grupy: A. z blizną trójpromienną - Triletes, B. bez blizny - Aletes i C. z blizną pojedynczą - Monoletes. W obrębie każdej z tych grup utworzył Ibrahim podgrupy oparte na morfologii ściany spor. Dla grupy A końcówka nazwy określającej typ morfologiczny była "i", np. Tuberculati-Sporites, Valvisi-Sporites dla grupy B końcówką było "a" - Punctata-Sporites, Zonala-Sporites Dla grupy C końcówka "o" Laevigato-Sporites, Zonalo-Sporites.
Hierarchiczne systemy klasyfikacji sporomorf karbońskich zostały rozbudowane przez Potoniego i Krempa (1954), którzy wprowadzili jako jednostki wyższego rzędu pojęcie "turma" (jest wiec anteturma jako jednostka najwyższa turma, subturma, infraturma, itp.). Ten typ klasyfikacji rozszerzono na zarodniki i ziarna pyłku roślin mezozoicznych (Dettman 1963).
Erdtman (1947) zaproponował morfograficzny system (NPC) dla klasyfikacji wszystkich spor i ziaren pyłku, zarówno kopalnych jak i współczesnych. W systemie tym sporomorfy zostały pogrupowane w oparciu o typ, liczbę i usytuowanie aparatów germinalnych (miejsc kiełkowania gametofitu). Takich grup wyróżnił Erdtman trzynaście, nie zaproponował jednak nadawania nazw gatunkowych i rodzajowych zgodnych z morfologią. W swoim podręczniku (1969) wyrażał się uszczypliwie o budowaniu nazw rodzajowych zawierających informacje o morfologii sporomorf.
Palinolodzy niemieccy (Potonié, Ratz i Thiergart w latach 1931-1938) dla określenia typu sporomorfy zastosowali nazwy Sporites i Pollenites. Przy kreowaniu gatunków, głównie pochodzących z osadów trzeciorzędowych, aby zaznaczyć ich możliwe pokrewieństwo z rodzajami roślin współczesnych, używali zapisu: np. Carya-Pollenites simplex R. Pot., Pterocarya-Pollenites stellatus (R. Pot.) Thiergart. W tym samym czasie palinolog amerykański (Wodehouse 1933) zaproponował dodanie przyrostka "pites" do botanicznego rdzenia w randze rodzaju (np. Caprifoliipites).
W roku 1950 Potonié, Thomson i Thiergart postanowili uporządkować ten typ nazewnictwa i ustanowili 10-punktowy projekt postępowania przy opracowaniu rozproszonych sporomorf trzeciorzędowych. (Potonié, Thomson, Thiergart 1950).
Przy monograficznym opracowaniu pyłku i zarodników należy, o ile to możliwe, umieszczać je w systemie naturalnym roślin.
(1) Istniejącą już nomenklaturę, obejmującą sporomorfy starsze (do starszego trzeciorzędu włącznie), należy zachować tak długo, dopóki nie da się zidentyfikować danej formy z odpowiednią rodziną, rodzajem, sekcją lub gatunkiem.
(2) Określenie typów pyłku i spor powinno być dokonywane z pełną świadomością, że mogą one obejmować wiele gatunków.
(3) Identyfikowanie typów powinno postępować od czwartorzędu do trzeciorzędu.
(4) Jeśli nie ma możliwości zupełnego zidentyfikowania danej sporomorfy ze współczesnym gatunkiem lub rodzajem, oznacza się ją z przyjętymi w paleobotanice regułami nomenklatury.
(5) Dodatek "typ" do nazwy rodziny oznacza, że możliwa jest także przynależność do innych rodzin. Należy wtedy do nazwy rodziny dodać wstawkę "oid" Jeśli jednak tylko jedna rodzina współczesna ma sporomorfy tego typu, to ani wstawka "oid" ani dodatek typ nie są potrzebne.
(6) Jeżeli rodzina jest oznaczona w sposób pewny, a rodzaj budzi wątpliwości, dodaje się do nazwy rodzajowej "typ" i wstawkę "oid". Jeśli rodzaj jest oznaczony z pewnością oba dodatki nie są potrzebne.
(7) Jeżeli chcemy przy dobrze oznaczonym rodzaju podać przypuszczalną przynależność do gatunku, to oba dodatki "typ" i "oid" umieszcza się przy nazwie gatunkowej.
(8) Jeśli używa się nazw pomocniczych (morfologicznych) dla rodzin, rodzajów lub gatunków, wstawka "oid" nie jest potrzebna, nie ma bowiem obawy pomyłki. Można jednak dodać do nazwy "typ".
(9) Dodatek "typ" nie jest potrzebny, jeśli jest zupełnie jasne, że istnieje tylko jeden rodzaj oznaczony nazwą pomocniczą.
Nadawanie nazw rodzajowych, traktowanych jako "pomocnicze", było dość ciekawą koncepcją palinologów w owym czasie. Zdawali oni sobie sprawę z możliwości późniejszej identyfikacji sporomorf z roślinami w naturalnym systemie klasyfikacji i liczyli się z możliwością zmiany pomocniczej nazwy rodzajowej (morfologicznej) na bardziej naturalną.
Powstała więc koncepcja rozdzielenia nazw rodzajowych na:
Genus - nazwa rodzajowa dla całej rośliny (przeważnie żyjącej).
Organ-genus - (rodzaj organowy), nazwa rodzajowa oparte na poszczególnych organach roślin z końcówką wskazującą na organ, na którego podstawie został ustanowiony.
Form-genus (rodzaj formowy) dla rozproszonych części roślin o nieznanym pokrewieństwie botanicznym.
Taki system tworzenia nazw rodzajowych był zapisany w Międzynarodowych Kodeksach Nomenklatury Botanicznej z Montrealu czy Seattle - przed 1975 rokiem.
Inny trend w nomenklaturze trzeciorzędowych sporomorf został oparty wyłącznie na cechach morfologicznych. Najbardziej znany system został opracowany przez Thomsona i Pfluga w 1953 roku. Według tego systemu pierwsza część nazwy rodzajowej informuje o ilości, typie i ułożeniu aparatów germinalnych (np. Triplano-, Tricolporo-), druga o typie organu (-sporites, -pollenites). Hierarchiczność tego systemy oparta jest również na morfologicznych jednostkach (Abteilung), bez wskazania pokrewieństwa botanicznego. Pokrewieństwo to jest uwzględnione jako jedna z informacji po opisie morfologicznych cech gatunku.
Palinolodzy zajmujący się trzeciorzędowymi sporomorfami i rozumiejący braki w możliwości wskazania pokrewieństw botanicznych już w nazwie rodzajowej zaproponowali powrót do początkowej praktyki palinologów niemieckich wprowadzając wskazanie na pokrewieństwo botaniczne w nazwie rodzaju. Takie propozycje przedstawili Potonié (1959-1971), Krutzsch (1959-1971), Nagy (1969, 1985), Planderova (1990) oraz autorzy polskiego opracowania sporomorf neogeńskich (Ziembińska et al. 1993).
Powrót do dawnej koncepcji "pomocniczych rodzajów" morfologicznych i zamiany ich na rodzaje organowe, wskazujące na pokrewieństwo botaniczne okazał się jednak niemożliwy w świetle nowych przepisów ICBN (1978 i 1988) w których zostały zlikwidowane inne "typy rodzajów", czyli rodzaj organowy i rodzaj formowy. Do wszystkich nazw rodzajowych - niezależnie od tego, czy jest to rodzaj organowy, czy "pomocniczy" formowy stosuje się prawo priorytetu. W wyniku ścisłych reguł ICBN (Międzynarodowego Kodeksu Nomenklatury Botanicznej) wszystkie nazwy rodzajowe utworzone po 1930 roku w sposób formalny (to znaczy mają wskazany typ, ilustrację i wystarczający opis) uważa się za ważnie opublikowane. W tej sytuacji dochodzi do paradoksalnych sytuacji, kiedy ewidentnie określony pyłek rodzaju Tsuga nazywa się Zonalapollenites, a zarodnik Sphagnum - Stereisporites. Stosowanie przepisów prawnych, dotyczących nomenklatury roślin jest niewątpliwie konieczne, ale nie uwzględnienie w nim specyfiki nomenklatury i taksonomii roślin kopalnych prowadzi do szeregu paradoksów.
Literatura
Chaloner, W.G. 1986. Reassembling the whole fossil plant, and naming it. Systematics Association Special Volume 31, 67-78.
Dettmen, M.E. 1963. Upper Mesosoic microfloras from South-Eastern Australia. Proceedings of the Royal Society of Victoria 77, 1-148.
Erdtman, E. 1947. Suggestions for the classification of fossil and recent pollen grains and spores. Svenska Botaniska Tidskrift 41, 104-114.
Greuter et al. 2000. International Code of Botanical Nomenclature. International Association
Krutzsch, W. 1959-1971. Atlas der mittel- und jungtertiären dispersen Sporen und Pollen - sowie der Mikroplanktonformen des nördlichen Mittereuropas.
Nagy, E. 1985. Sporomorphs of the Neogene in Hungary. Geologia Hungarica, series Paleontology 47.
Planderova, E. 1990. Miocene microflora of Slovak Central Paratethys and its biostratigraphical significance. Dionyz Stur Institut Geologii, Bratislava.
Potonié, R. 1956-1971. Synopsis der Gattungen der Sporae dispersae. Geologisches Jahrbuch, Beiheft
Potonié, R. & Kremp, G. 1954. Die Gattungen der paläozoischen Sporae disperse und ihre Stratigraphie. Geologisches Jahrbuch 69, 111-194.
Potonié R., Thomson, P.W., & Thiergart F. 1950. Zur Nomenklatur und Klassifikation der neogenen Sporomorphae (Pollen und Sporen). Geologisches Jahrbuch 65, 35-70.
Traverse, A. 1996. Nomenclature and taxonomy: systematics. A rose by any other name would by very confusing. In J. Jansonius & D.C. McGregor (eds) Palynology: Principles and Applications. Vol. 1., 11-28. AASP, Salt Lake City.
Wodehouse, R.P. 1933. Tertiary pollen II. The oil shales of the Green River Formation. Bulletin of the Torrey Botanical Club 60, 479-524.
Ziembińska et al. 1993. Taxonomical revision of selected pollen and spores taxa from Neogene deposits. In L. Stuchlik (ed.) Neogene pollen flora of Central Europe, part 1. Acta Palaeobotanica, Suplement 1, 5-30.
Problemy z doborem typów opisowych w paleontologii kręgowców
MARIUSZ LUBKA
Uniwersytet Wrocławski
Naczelnym dokumentem definiującym pojęcie typu opisowego oraz jego stosowanie w taksonomii zoologicznej jest Międzynarodowy Kodeks Nomenklatury Zoologicznej. Nie bez przyczyny Kodeks poświęca pojęciu typu wiele stron. Jest bowiem typ obiektywnym i niezmiennym onomatoforem nazwy gatunku do którego należy, swoistym wzorcem odniesienia w przypadku sporu nad zaliczeniem danego okazu. Bez względu na to, jak zmienia się pojmowanie taksonu, jego typ pozostaje ten sam. Jest oczywiste, iż gatunek złożony z licznych, często znacznie różniących się między sobą populacji, nie może zostać sprowadzony do pojedynczego okazu, typ ma zatem znaczenie czysto formalne. Ustanawiający go badacz winien dbać o to, by był typowy dla reprezentowanego gatunku. Aby ustabilizować nomenklaturę pojęcie typu rozszerzono na kategorie ponadgatunkowe. Tak więc typem rodzaju jest gatunek typowy, typem rodziny rodzaj nominalny, itd.
Opis nowego gatunku opiera się najczęściej na serii okazów. Zgodnie z nakazem Kodeksu autor powinien wybrać spośród nich jeden jedyny, który stanie się nośnikiem nazwy - holotyp. Rzadko, szczególnie w paleontologii, zdarza się, że holotyp posiada wszystkie cechy diagnostyczne gatunku, o których istnieniu wiemy z innych okazów. Jeśli wymienia się w oryginalnym opisie gatunku okazy inne niż holotyp, oznaczone są one jako paratypy. Paratypów może być dowolnie dużo, w praktyce jednak z długich serii wybiera się ich jedynie kilka. Kodeks nie wyróżnia tu innych niż wspomniane typów. Zwyczajowo jednak wymienia się też allotyp (egzemplarz płci przeciwnej do holotypu) lub/i pedotyp (okaz larwalny, bardzo często wskazywany np. w entomologii). Szczególnie w paleontologii rozpowszechnione jest nieformalne pojęcie hipodygmatu określające zbiór okazów przypisanych do jednego gatunku. Syntypy to wszystkie okazy typowe, jakie stały się podstawą wyznaczenia gatunku.
Autorzy osiemnastowieczni i dziewiętnastowieczni nie wyznaczali okazów typowych przy opisie nowego gatunku. Wszystkie okazy, które autor miał do dyspozycji jako podstawę nadania nazwy mają równą wartość nomenklatoryczną. Badacz rewidujący dany gatunek może spośród nich wybrać jeden okaz będący odpowiednikiem holotypu - lektotyp. W końcu zdarza się też, że holotyp czy lektotyp ulegają zniszczeniu lub zagubieniu. W wyjątkowych przypadkach, kiedy zachodzi konieczność rozstrzygnięcia ważkich problemów taksonomicznych Kodeks dopuszcza wyznaczenie neotypu. Najistotniejsze przy wyznaczaniu neotypu jest, by pochodził z obszaru położonego jak najbliżej miejsca pochodzenia holotypu.
Zasady doboru okazów typowych dokonywanego przez paleontologów nie różnią się od doboru czynionego przez "neontologów". Tyle, że oparty jest on na materiale z natury często fragmentarycznym i źle zachowanym. Zdarza się, że nowe nazwy gatunkowe nadawane są niemal każdej nowo odkrytej skamieniałości kopalnego kręgowca. Efektem są długie listy synonimów. Historia nomenklatoryczna niektórych taksonów jest bardzo niejasna i skomplikowana.
Problem Parasuchus
Tego typu problemy ujawniają się przy wyborze formalnie poprawnej nazwy rodzajowej fitozaura z Krasiejowa. Stan zachowania wydobytych szczątków jest bardzo dobry i znalezienie cech diagnostycznym nie było trudne. Do tego rodzaju prymitywnego karnijskiego fitozaura używa się jednak dwóch nazw: Paleorhinus i Parasuchus. Są one oczywiście synonimami, wydawałoby się zatem, iż wystarczy ustalić który z nich jest starszy i wykorzystując jedno z podstawowych praw nomenklatury jakim jest zasada priorytetu ostatecznie ustalić nazwę. Sprawa jest jednak dalece bardziej skomplikowana.
Historia statusu nomenklatorycznego Parasuchus hislopi i jego jednostek ponadgatunkowych jest zagmatwana od półtora wieku. Huxley (1870) jako pierwszy użył nazwy Parasuchus w swej liście fauny triasu, nie podając jednak żadnej diagnozy. Nazwa pozostawała nomen nudum (nazwa taksonomiczna nie poparta opisem ani nie przywiązana do okazu typowego) do roku 1885, kiedy to Lydekker opisał gatunek fitozaura Parasuchus hislopi, opierając się na serii typowej kości, tarczek i zębów pochodzących z formacji Maleri z doliny Pranhita-Godavari w Indiach. P. hislopi stał się jednocześnie gatunkiem typowym rodzaju Parasuchus, i typem wydzielonej przez Lydekkera nowej rodziny Parasuchidae. Po pewnym czasie okazało się niestety, iż wśród syntypów P. hislopi znajdowały się szczątki rynchozaura [basicranium Paradapedon huxleyi (Lydekker, 1881); von Huene 1940]. Status Parasuchus pozostawał problematyczny przez następnych wiele lat, rodzaj był uznawany za "wątpliwy", czasem nawet klasyfikowany jako rynchozaur (Hoffstetter 1955). Później Colbert (1958) zaliczył cały materiał indyjski do gatunku Phytosaurus malariensis, ale Gregory (1962) uważał, że nie ma on jednak cech diagnostycznych.
Oryginalny materiał Lydekkera to źle zachowane fragmenty kości nierozpoznawalne na poziomie rodzajowym (Long & Murry 1995). Zimą 1965-66 w dolinie Pranhita-Godavari odkryte zostały jednak dwa, świetnie zachowane, artykułowane szkielety fitozaurów. Opracowujący je S. Chatterjee zajął w 1974 stanowisko, iż materiał Lydekkera jest jednak diagnostyczny. Chcąc uprawomocnić nazwę P. hislopi, wyselekcjonował spośród oryginalnych okazów Lydekkera fragment przodu pyska fitozaura (Indian Museum, Calcutta, GSI H20/11) ustanawiając lektotyp. Uznał, że skoro basicranium rynchozaura (jeden z syntypów Lydekkera) nie jest ani holotypem Parasuchus hislopi, ani też lektotypem Paradapedon huxleyi można uniknąć zaniechania nazwy Parasuchus hislopi i powrócić do pierwotnej intencji Lydekkera (tzn., że nazwa należy do fitozaura). Umożliwić to miało zachowanie stabilności nomenklatury. Konsekwentnie w 1978 zaliczył szkielety z formacji Maleri do gatunku P. hislopi. Lektotyp, zgodnie z rozporządzeniem Kodeksu (Art.74.1), musi zostać wybrany spośród oryginalnych okazów typowych (paratypów syntypu) na podstawie których opisano gatunek. Jeśli okazałoby się, że okaz określony jako lektotyp nie należał do okazów typowych, straci swój status (Art. 74.2). Chatterjee, chcąc uprawomocnić nazwę, musiał więc wskazać jako wzorzec gatunku jeden ze źle zachowanych, nie wykazujących cech diagnostycznych okazów Lyddekera, mimo iż był już wtedy w posiadaniu dwóch doskonale zachowanych szkieletów.
Autor trafnie zauważył, że indyjski Parasuchus Lydekker, 1885 (w jego rozumieniu) oraz północnoamerykański i europejski Paleorhinus Williston, 1904 są tak podobne, że nie sposób ich rozróżnić na poziomie rodzajowym. Szkielety z Maleri były bez wątpienia kongeneryczne z Paleorhinus bransoni Williston, 1904 z formacji Popo Agie z Wyoming. Ponieważ Chatterjee uważał okazy Lydekkera z 1885 oraz kompletne, opisywane przez siebie szkielety występujące w tej samej formacji za konspecyficzne (tym samym wyrażając zdanie, że gatunek Lydekkera jest prawidłowy), wyraził pogląd, iż rodzaj Willistona, ustanowiony w 1904 r. na podstawie dobrze zachowanej, prawie kompletnej czaszki (CFMNH UR632) jest młodszym synonimem Parasuchus. Dodatkowo zasugerował ustanowienie dwóch podrodzajów (Parasuchus i Paleorhinus) w obrębie rodzaju Parasuchus, odzwierciedlające geograficzne oddalenie bliskich sobie pod względem morfologicznym gatunków. Już w momencie ogłoszenia odkrycia w Indiach artykułowanych szkieletów pojawiły się głosy, iż lektotyp Parasuchus hislopi jest zbyt fragmentaryczny i niediagnostyczny. W związku z tym niektórzy autorzy sugerowali, by nazwę rodzaju Parasuchus (którego gatunek P. hislopi jest gatunkiem typowym) zastąpić nazwą Paleorhinus.
Gdyby lektotyp Parasuchus był w lepszym stanie (zawierał choćby jedną cechę diagnostyczną), a w formacji Maleri nie występowałyby inne fitozaury, można by uznać, iż szkielety Chatterjee'go należą do prawidłowego rodzaju Parasuchus, która to nazwa miałaby priorytet przed Paleorhinus. Jednakoż von Huene (1940) opisał z doliny Pranhita-Godavari fragment bardzo dużego rostrum fitozaura (GSI No. 16691) i skojarzony z nim kawałek żuchwy (GSI No. 16693) nadając mu nazwę "aff. Brachysuchus" malariensis. Materiał nie jest rozpoznawalny na poziomie rodzajowym, z pewnością jednak nie jest kongeneryczny z Paleorhinus (a tym samym Parasuchus) i należy do bardziej zaawansowanego fitozaura z wysokim rostrum i przesuniętymi dalej ku tyłowi nozdrzami (Long & Murry 1995 za Gregory 1962; Colbert 1958). Chatterjee jednak błędnie uznał, że materiał von Huene'go reprezentuje ten sam takson, co jego dwa szkielety.
Zważywszy na niekompletność materiału Lydekkera i obecność więcej niż jednego gatunku fitozaura w formacji Maleri, Long i Murray (1995) zaproponowali uznać oryginalny materiał Parasuchus hislopi jako przedstawiciela fitozaurów o nieustalonych pokrewieństwach (Parasuchia ident.), zaś artykułowane szkielety Chatterjee'ego zaliczyć do rodzaju Paleorhinus (P. sp.). Okaz "aff Brachysuchus" malariensis został przez tych autorów również uznany za nomen dubium (nazwa taksonu, którego gatunek typowy nie posiada cech diagnostycznych wystarczających do odróżnienia go od podobnych okazów innych gatunków) w obrębie Parasuchia, różny od Paleorhinus i zapewne reprezentujący linię rutiodontów. Moim zdaniem to dobre rozwiązanie. Tym bardziej, że Paleorhinus - dobrze poznany już w początkach XX wieku - wydaje się być silnie zakorzeniony w literaturze i świadomości naukowców.
Historia się na tym jednak nie kończy. W marcu 2001 roku Chaterjee złożył do Międzynarodowej Komisji Nomenklatury Zoologicznej wniosek, którego celem było wyznaczenie neotypu dla Parasuchus hislopi Lydekker, 1885. Aby utrzymać stabilność używania nazwy i w zgodzie z Artykułem 75.5. Kodeksu zaproponował odrzucić lektotyp a kompletny artykułowany szkielet z Maleri wyznaczyć jako neotyp. Trudno przewidzieć decyzję Komisji. Międzynarodowy Kodeks Nomenklatury Zoologicznej wyraźnie określa warunki w których można wyznaczyć neotyp: dzieje się to tylko wtedy, gdy istnieje pewność, że holotyp lub lektotyp zaginął lub został zniszczony, a przy tym wystąpiła konieczność rozwiązania ważnego problemu natury nomenlkatorycznej. Jednak właśnie Art.75.5. dopuszcza ustanowienie neotypu, kiedy gatunku nie można zidentyfikować na podstawie istniejącego okazu typowego (holotypu bądź lektotypu), tzn. kiedy nazwa jest nomen dubium.
Literatura
Chaterjee, S. 1974. A rhynchosaur from the Upper Triassic Maleri Formation of India. Philosophical Transactions of the Royal Society of London B 267, 209-261.
Chatterjee, S. 1978. A primitive parasuchid (phytosaur) reptile from the Upper Triassic Maleri Formation of India. Palaeontology 21, 83-127.
Chatterjee, S. 2001. Parasuchus hislopi Lydekker, 1885 (Reptilia, Archosauria): proposed replacement of the lectotype by a neotype. Bulletin of Zoological Nomenclature 58 (1), case 3165.
International Commission on Zoological Nomenclature. 1999. International Code of Zoological Nomenclature, 4th Edition. 1-126. The International Trust for Zoological Nomenclature, London.
Long, R.A. & Murry, P.A. 1995. Late Triassic (Carnian and Norian) tetrapods from the Southwestern United States. Bulletin of the New Mexico Museum of Natural History and Science 4, 1-254.
Nomenklatoryczny i taksonomiczny status podgatunku w paleozoologii kręgowców
TOMASZ SULEJ
Instytut Paleobiologii PAN
Od zarania biologii istniała tendencja do nadmiernego mnożenia liczby gatunków. Choć już Darwin rozważał istnienie ras geograficznych, nawet niewielkie różnice między populacjami uznawano za cechy gatunkowe. Dopiero wprowadzenie biologicznej koncepcji gatunku umożliwiło zobiektywizowanie kryteriów wydzielania gatunków. Pojawił się wraz z tym problem populacji, które wykazują odmienność morfologiczną czy anatomiczną, ale mogą się krzyżować. Rozdziela je tylko przestrzeń. Dla takich "niepełnych" gatunków, czyli ras geograficznych stosuję się rangę podgatunku.
Mayr (1974) definiuje podgatunek jako zbiór fenotypowo podobnych populacji danego gatunku, zamieszkujący geograficznie wydzieloną część areału gatunku i różniący się taksonomicznie od innych populacji tego gatunku. Różnice taksonomiczną rozumie jako dostatecznie wyraźne diagnostyczne cechy morfologiczne. Dziś uwzględnia się też różnice genetyczne. Z definicji zasięgi geograficzne dwóch podgatunków nie mogą się na siebie nakładać.
| 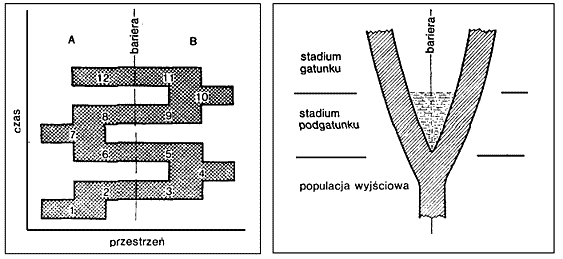 | Ryc. 1. Geografia i czas w tworzeniu gatunków i podgatunków (wg Mayr 1974). |
W paleontologii nie jest łatwo zastosować pojęcie podgatunku rozumianego jako rasa geograficzna. Trudno bowiem określić, na ile różnice między profilami geologicznymi są przejawem różnic czasowych, a na ile różnic środowiskowych (Rys. 1). Wymaga to precyzyjnej korelacji wiekowej i pełnej dokumentacji w różnych profilach geologicznych. Znacznie częściej wyróżnia się natomiast podgatunki temporalne, rozumiane jako wycinki ciągu przemian ewolucyjnych o odmienności mniejszej od tej, którą uznaje się za właściwą dla rangi gatunku. W każdym momencie swojego istnienia (przekroju czasowym) podgatunek temporalny był pełnym gatunkiem biologicznym. Pomiędzy podgatunkiem rozumianym jako obiektywnie odrębna rasa geograficzna, a podgatunkiem rozumianym jako arbitralnie wydzielony wycinek ewolucji jest więc fundamentalna odmienność. Były próby zaznaczenia tego w zapisie nazw, ale nie upowszechniły się, bowiem w biologii dzisiejszej dominuje pogląd o nagłym pojawianiu się gatunków i obiektywności ich granic również w czasie geologicznym.
Podgatunek dzisiejszych kręgowców
Obecnie dostępne techniki sekwencjonowania genów umożliwiają szybkie porównywanie genomu dużej liczby osobników. Metody te coraz szerzej stosuje się także w biogeografii. Przykładem może być opracowanie taksonomii morszczuka przez Grant & Leslie (2001), którzy porównali sekwencje 11 enzymów u większości gatunków morszczuka z danymi morfologicznymi i paleontologicznymi.
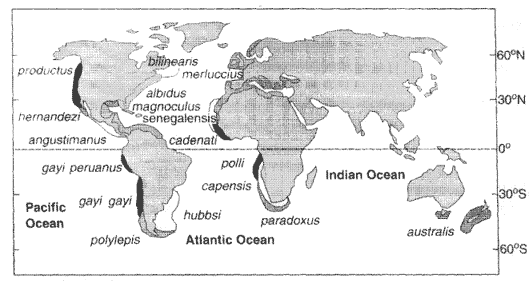 | Ryc. 2. Geograficzne rozmieszczenie gatunków morszczuka Merluccius. |
Rozpatrując uzyskane wyniki w kontekście zasięgów geograficznych badanych populacji i barier między nimi, uznali dwa wyróżniane do tej pory gatunki za podgatunki, zaś wątpliwe gatunki okazały się rzeczywistymi. W pierwszym przypadku podobieństwo genetyczne między gatunkami Merluccius cadenati i M. polli oraz M. polylepis i M. australis jest na tyle duże, że wyklucza różnice gatunkowe, jednocześnie badania wskazują na wyraźną odrębność między obiema populacjami, a ponieważ są one oddzielone barierą w postaci nie mieszających się wód, uznano je za podgatunki.
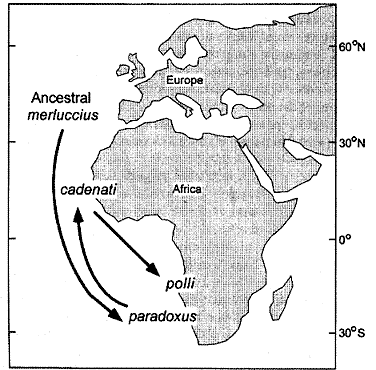 | Ryc. 3. Model migracji i ewolucji morszczuka Merluccius. M. paradoxus datuje się na 10 Ma. Z niego prawdopodobnie 5-6 Ma wyewoluował M. cadenati, w którego obrębie 1 Ma nastąpiło rozdzielenie na dwa podgatunki. Jeden z nich, żyjący na południu - M. polli, do tej pory uważany był za gatunek, jednak bardzo duże podobieństwo jego genomu do M. cadenati przeczy takiemu przyporządkowaniu. |
Drzewa rodowe oparte na danych molekularnych wymagają kalibracji przy pomocy danych paleontologicznych. Wymaga to sformułowania hipotez dotyczących zarówno materiału dzisiejszego, jak kopalnego. Przykład badań Grant & Leslie (2001) pokazuje, że połączenie danych genetycznych i paleontologicznych umożliwia udokumentowanie filogenezy i zobiektywizowanie pojęć gatunku i podgatunku.
Europejskie podgatunki triasowego płaza Metoposaurus
W Europie szczątki metopozaurów znane są z kilku stanowisk w obrębie Basenu Germańskiego. Najstarsze pochodzą z piaskowca trzcinowego (Schilfsandstain). W osadach tych w okolicach Stuttgartu znaleziono czaszki, kości pasa barkowego, a także artykułowany fragment szkieletu. To właśnie stamtąd zostały opisane pierwsze metopozaury, w tym holotyp Metoposaurus diagnosticus. Pojedyncze czaszki zaliczane do tego gatunku pochodzą także z osadów leżących nad Schilfsandstein, tj. Rote Wand, najmłodsze z Kieselsandstain. W zalegających między nimi warstwach z Lehrberg znalezione zostały międzyobojczyki. Prawdopodobnie tego samego wieku są liczne okazy, w tym także czaszki, z Krasiejowa na śląsku Opolskim (Dzik et al. 2000). Metopozaury z wszystkich tych stanowisk mają wiele cech wspólnych, należą więc prawdopodobnie do jednego gatunku Metoposaurus diagnosticus. Monotypowy rodzaj Metoposaurus charakteryzuje się kością łzową, która tworzy krawędź oczodołów oraz bardzo małą powierzchnią z izometrycznym urzeźbieniem na międzyobojczyku. Pomiary kości czaszek z różnych stanowisk ujawniły jednak istotne różnice w obrębie gatunku.
Zróżnicowanie wewnątrzgatunkowe Metoposaurus
Seria 14 dotąd odpreparowanych czaszek z Krasiejowa jest najliczniejszą próbką z populacji metopozaurów Basenu Germańskiego. Z Schilfsandstein pochodzą trzy czaszki, po jednej z Rote Wand i z Kieselsandstein. Ponieważ są one w większości niekompletne, porównanie ich proporcji nie jest możliwe. Pozostają badania pojedynczych kości. Już Branson i Mehl (1929) zauważyli pewne różnice między gatunkami metopozaurów w obrębie kształtu kości ciemieniowej. Proporcje te uwidacznia odległość między środkiem otworu ciemieniowego a najdalej do przodu wysuniętą krawędzią kości ciemieniowej (U) oraz kąt między lewym i prawym szwem kości ciemieniowej i nadciemieniowej (?). Szerokość czaszek może służyć jako przybliżony wskaźnik zawansowania wzrostu (Ryc. 4 i 5).
Są wyraźne, statystycznie istotne różnice w proporcjach kości ciemieniowej między populacją z Schilfsandstein i z Krasiejowa. Czaszki z Krasiejowa mają wyraźnie krótszą przednią część oraz większy kąt alfa.
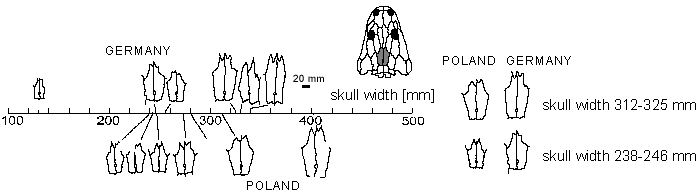 |
Ryc. 4. Obrysy kości ciemieniowych metopozaurów z Wirtembergii i z Polski, ułożone według szerokości całej czaszki. Z prawej strony kości ciemieniowe czaszek o tej samej szerokości przedstawione - wyraźnie widać różnice w długości kości ciemieniowej. |
Biologiczny sens zmian kształtu kości ciemieniowej. - Dzięki urzeźbieniu powierzchni kości, bardzo wyraźnie widać istnienie stref szybszego wzrostu czaszki metopozaurów. Można u nich wyróżnić dwa rodzaje ornamentacji dachu czaszki: izometryczne zagłębienia oznaczają strefę rosnącą stosunkowo powoli, natomiast wydłużone rowki są oznaką intensywnego rozwoju kości, powstają bowiem przez wydłużenie izometrycznych dołków.
Strefa intensywnego wzrostu biegnąca w poprzek czaszki znajduje się za oczodołami, na wysokości przedniej części kości ciemieniowej. Zdecydowanie wolniej przyrastała strefa między oczodołami i nozdrzami. Te proporcje obrazują w pewnym stopniu ewolucję w linii prowadzącej do metopozaurów. Polegała na przesuwaniu oczodołów do przodu czaszki (Schoch & Milner 2000). Było to przejawem pedomorfozy, bowiem oczodoły znajdują się z przodu czaszki u młodych temnospondyli (Warren & Hutchinson 1988). Ich usytuowanie u form dorosłych jest wynikiem różnego tempa wzrostu wspomnianych stref czaszki. I tak np. u kapitozaurów mających oczodoły z tyłu czaszki, wszystkie znajdujące się za nimi kości mają rzeźbę tylko w postaci izometrycznych zagłębień, natomiast długimi rowkami pokryte są wydłużone kości strefy przedoczodołowej. Linia ewolucyjna prowadząca do metopozaurów oddzieliła się od linii kapitozaurów już we wczesnym triasie. U kolejnych jej przedstawicieli obserwuje się stopniowe przesuwanie oczodołów do przodu czaszki. Prawdopodobnie było to związane z przystosowaniem do bardziej wodnego, niż kapitozaury, trybu życia. Metopozaury są zakończeniem tej linii i mają najbardziej do przodu wysunięte oczodoły.
Próba interpretacji taksonomicznej
Ze względu na duże podobieństwo anatomii w obrębie metopozaurów i trudności z datowaniem poszczególnych stanowisk rozpoznanie ich ewolucji nastręcza duże trudności. Niewątpliwie istotne są różnice w proporcjach kości ciemieniowej pomiędzy populacjami z Schilfsandstein i warstw z Lehrberg. Prawdopodobnie reprezentują one różne podgatunki Metoposaurus diagnosticus. Nie jest jednak jasne znaczenie różnic w budowie czaszki między populacjami z czterech kolejnych horyzontów czasowych z Basenu Germańskiego. Jednoznaczne określenie, czy były to podgatunki temporalne czy geograficzne jest przez to niemożliwe. Różne cechy jednej czaszki pochodzącej z Kieselsandstein z Wirtembergii, a więc najmłodszej, wspierają przeciwstawne hipotezy.
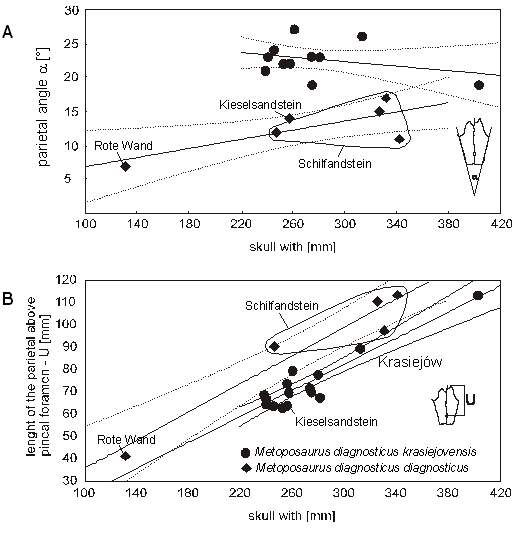 | Ryc. 5. Zmiany proporcji kości ciemieniowej w ontogenezie.
A. Niejasna zależność wartości kąta parietale od szerokości czaszki. Prawdopodobnie odmienności regresji są wynikiem zbyt małych próbek, a kąt ten nie zmienia się w ontogenezie.
B. Różnice w wydłużaniu kości ciemieniowej u podgatunków Metoposaurus diagnosticus. |
Interpretacja geograficzna. - Czaszka z Kieselsandstein wraz z pozostałymi czaszkami z niemieckiej części Basenu Germańskiego odróżnia się od serii z Krasiejowa małą wartością kąta parietale. Sugeruje to istnienie dwóch ras geograficznych. Jena z nich mogła występować w zachodniej części basenu, obejmując wszystkie niemieckie metopozaury charakteryzujące się małą wartością ?, druga we wschodniej reprezentowanej przez okazy z Krasiejowa. Hipotezę tę wzmacnia występowanie bariery geograficznej w postaci pasma dzisiejszych Sudetów, które w trakcie obniżenia poziomu morza (Schilfsandstein) mogły rozdzielić wschodnią i zachodnią część basenu, wprowadzając czynnik izolacji geograficznej umożliwiający rozbieżną ewolucję.
Interpretacja temporalna. - Pod względem wydłużenia kości ciemieniowej czaszka z Kieselsandstein jest jednak podobna do populacji z Krasiejowa. Jeśli założyć, że tworzyły jedną populację, byłaby ona młodsza od populacji z Schilfsanstein i Rote Wand. Tym samym wskazywałoby to na istnienie jednego ciągu ewolucyjnego populacji zasiedlających cały Basen Germański. Populacja z Schilfsandstein, populacja z Krasiejowa i populacja z Kieselsandstein to tylko kolejne stadia przemian. Odmienności morfologiczne wystarczają do wydzielenia dwóch łatwo rozpoznawalnych stadiów - podgatunków temporalnych. Podgatunek starszy miałby dłuższe parietale, natomiast ewolucyjnie późniejszy krótsze. Ewolucja polegałaby na zmianie rozwoju czaszki w ontogenezie. Wyrażałoby się to wolniejszym rozwojem kości ciemieniowej na długość i jednocześnie poszerzaniem się jej na boki. Mogło to być wynikiem zmian zachodzących w całej czaszce, jednakże ze względu na niekompletność okazów trudno to przedstawić ilościowo. Problem ten dotyczy zresztą całego zagadnienia, rozważanego na podstawie tylko jednej czaszki, co osłabia wiarygodność przedstawianych hipotez.
Wniosek. - Ze względu na bardzo duże podobieństwo długości kości ciemieniowej okazu z Kieselsandstein do jednorodnej populacji z warstw z Lehrberg oraz oszczędność metodologiczną (nie trzeba wprowadzać bariery geograficznej), za właściwszą należy uznać hipotezę proponującą wprowadzenie podgatunku temporalnego. Proponuje się więc wprowadzenie podgatunku Metoposaurus diagnosticus krasiejovensis dla populacji Metoposaurus diagnosticus występującej w osadach warstw z Lehrberg, Krasiejowa i Kieselsandstein. Okazy pochodzące z osadów Schilsandstein i Rote Wand należałyby do Metoposaurus diagnosticus diagnosticus.
Literatura
Branson, E. B. & Mehl, M. G. 1929. Triassic Amphibians from the Rocky Mountain Region. University of Missouri Studies 4, 155-239.
Dzik, J., Sulej, T., Kaim, A. & Niedźwiedzki, R. 2000. Późnotriasowe cmentarzysko kręgowców lądowych w Krasiejowie na śląsku Opolskim. Przegląd Geologiczny 48, 226-235.
Grant, W. S. & Leslie R. W. 2001. Inter-ocean dispersal is an important mechanism in the zoogeography of hakes (Pisces: Merluccius spp.). Journal of Biogeography 28, 699-721.
Mayr, E. 1974. Podstawy systematyki zwierząt. PWN, Warszawa
Schoch, R. R. & Milner, A. R. 2000. Stereospondyli. Handbuch der Paläoherpetologie teil 3B, pp.165.
Warren, A. A. & Hutchinson, N. 1988. A new capitosaurid amphibian from the early Triassic of Queensland, and the ontogeny of the capitosaur skull. Palaeontology 31, 857-876.
Nomenklatoryczne konsekwencje typologicznej i populacyjnej koncepcji gatunku w paleozoologii bezkręgowców
MICHAŁ ZŁOTNIK
Uniwersytet Warszawski
Oparta na skamieniałościach identyfikacja gatunków kopalnych stanowi fundament paleontologii, będąc dla wielu przedstawicieli tej dyscypliny naukowej celem samym w sobie, dla innych zaś zaledwie wstępem do bardziej wyrafinowanych analiz. Podjęcie decyzji o tym czy dana skamieniałość reprezentuje ten bądź inny gatunek kopalny nieuchronnie pociąga za sobą konsekwencje nomenklatoryczne, gdyż wiąże się z przypisaniem skamieniałości właściwej jej nazwy gatunkowej. Identyfikowanie skamieniałości (zwane w żargonie paleontologów ich oznaczaniem) jest czynnością wysoce subiektywną, gdyż trafność podejmowanych decyzji w znacznym stopniu zależy od kompetencji, doświadczenia, i intuicji badacza, w szczególności zaś od jego sposobu pojmowania gatunku. Ze względu na ten ostatni aspekt paleontologów można zasadniczo podzielić na zwolenników jednej z dwu koncepcji gatunku: ujęcia typologicznego bądź populacyjnego.
Ujęcie typologiczne zakłada, iż gatunek posiada pewne cechy absolutnie stałe, zaś wzorcem tych cech jest typowy okaz gatunku zwany holotypem. W swojej skrajnej wersji pozwala do danego gatunku zaliczyć wyłącznie okazy identyczne z holotypem. Holotyp pełni więc w niej funkcję podobną jak prototyp w świecie urządzeń będących wytworem człowieka.
Ujęcie populacyjne stoi w opozycji do typologicznego, gdyż eksponuje różnice wewnątrzgatunkowe i dopuszcza zaliczenie do jednego gatunku okazów nie tylko różniących się pomiędzy sobą, ale i różnych od holotypu. Holotyp odgrywa tu więc znacznie mniejszą rolę - jest przede wszystkim nośnikiem nazwy gatunku, w znacznie mniejszym stopniu zaś jego cech, które z założenia mogą być zmienne.
Ujęcie typologiczne jest sprzeczne z biologiczną koncepcją gatunku. Dane empiryczne wskazują bowiem w sposób jednoznaczny, iż organizmy należące do jednego gatunku biologicznego mogą pod względem wielu cech wykazywać daleko idące różnice, czego najlepszym a zarazem najbliższym nam przykładem może być nasz własny gatunek - Homo sapiens. Wiadomo o tym już od połowy XIX wieku. Pomimo, że ujęcie typologiczne jest ewidentnie błędne i powinna zostać już dawno zarzucone, jego duch przewija się w pracach dzisiejszych badaczy.
Typologiczne i populacyjne podejście do gatunku na przykładzie rodziny Nassaridae
Doskonałych przykładów nomenklatorycznych konsekwencji obu sposobów myślenia dostarcza historia wydzielania gatunków w obrębie rodziny Nassaridae - grupy przodoskrzelnych ślimaków, których muszle znajdowane są w licznych stanowiskach miocenu Europy.
Hinia styriaca. - Jednym z gatunków nassaridów, licznie reprezentowanym w iłach korytnickich oraz w innych osadach zdeponowanych w obrębie licznych basenów Paratetydy jest Hinia styriaca. Wysmukłość muszli tego gatunku wyrażona poprzez stosunek jej szerokości do wysokości (W/H) waha się u wyrośniętych okazów pochodzących z basenu Korytnicy w dość szerokim zakresie (0,46 - 0,60). Zmienność tej cechy ma charakter ciągły i unimodalny, przy czym okazy najbardziej wysmukłe i najbardziej pękate występują najrzadziej, dominują zaś okazy o wysmukłości pośredniej. Jest to rozkład zbliżony do normalnego i wskazuje na przynależność wszystkich okazów do jednej populacji biologicznej.
Rzecz interpretowana była zgoła inaczej w opracowaniach zwolenników typologicznego ujęcia gatunku. Hilber w roku 1879 uznał, że okazy wysmukłe reprezentują odmienny gatunek kopalny i nadał mu nazwę Hinia intersulcata. Niewątpliwie typologiczne stanowisko zajęli także badacze późniejsi, którzy zidentyfikowali ów gatunek z obszaru Włoch (Venzo & Pelosio 1963), Węgier (Kókay 1966) oraz Basenu Wiedeńskiego (Hoernes & Auinger 1882) pod zaproponowaną przez Hilbera nazwą. Okazy opisane i ilustrowane przez cytowanych autorów mieszczą się w spektrum zmienności korytnickich przedstawicieli Hinia styriaca. O ich typologicznej postawie świadczy to, iż dwu spośród nich rozpoznało w badanym przez siebie materiale także gatunek Hinia styriaca. Jest więc niemal pewne, iż autorzy ci posiadali wiele okazów o pośredniej wysmukłości, jednak ich przywiązanie do idei holotypu jako wzorca spowodowało, iż uznali gatunek wydzielony przez Hilbera. Przykład powyższy dotyczył zmienności osobników znajdujących się na podobnym stadium rozwojowym, gdyż wszystkie rozpatrywane okazy miały 5 skrętów teleokonchy (części muszli tworzonej przez ślimaka po przejściu metamorfozy).
Hinia daciae. - U nassaridów jest wyraźna tendencja do wysmuklania muszli w trakcie ontogenezy. Tendencję tę doskonale ilustrują muszle korytnickiego nassarida Hinia daciae reprezentujące kolejne stadia rozwojowe (3, 4 i 5 skrętów teleokonchy). Na diagramie ilustrującym zmienność H. daciae jawi się to jako stopniowe przesuwanie zakresu zmienności W/H w kierunku wartości niższych (Ryc. 2). W populacyjnym ujęciu gatunku wszystkie ilustrowane okazy reprezentują jeden gatunek - H. daciae, pomimo, iż różnią się pomiędzy sobą oraz są różne od holotypu tego gatunku. Holotyp pochodzący z innej lokalizacji jest bowiem okazem szczególnie wyrośniętym, posiadającym blisko 6 skrętów teleokonchy. Zgodnie z obserwowaną u nassaridów tendencją ma prawo być szczególnie wysmukły.
Badacze reprezentujący koncepcję typologiczną mieli na ten temat pogląd odmienny. Boettger (1906) wyróżnił odmianę z miocenu Transylwanii charakteryzującą się bardziej pękatą muszlą (Hinia daciae var. rejecta). Nie uwzględnił jednak tego, iż odnosi się do okazów mających zaledwie 3 skręty teleokonchy, podczas gdy holotyp gatunku blisko 6, były to więc okazy znajdujące się na zupełnie odmiennym stadium rozwojowym. Nic więc dziwnego, iż okaz uważany przez Boettgera za odmianę omawianego gatunku mieści się w spektrum zmienności okazów H. daciae z basenu Korytnicy.
Sphaeronassa dujardini i S. schoenni.- Typologiczne pojmowanie gatunku daje plon w postaci nowych taksonów, przede wszystkim tam, gdzie mamy do czynienia z gatunkami charakteryzującymi się szczególnie dużą zmiennością. Odpowiedniego przykładu dostarczają dwa kolejne gatunki nassaridów korytnickich: Sphaeronassa dujardini i S. schoenni. Oba gatunki charakteryzują się stosunkowo zmienną (a) wysmukłością muszli (W/H) i (b) względną wysokością ostatniego zwoju (mierzoną względem wysokości muszli Hlw/H). Ponadto wyrośnięte formy S. schoenni mają mniej lub bardziej wydatne osiowe fałdy na ostatnim zwoju. Ponieważ zakres zmienności cech metrycznych obu gatunków częściowo się pokrywa, obecność (lub brak) owych fałdów jest jedyną cechą dobrze oddzielającą oba gatunki. W oparciu o okazy pochodzące z basenu Wiedeńskiego Beer-Bistricky (1958) podzieliła S. dujardini na 3 podgatunki (opisane pod inną nazwą rodzajową): Hinia dujardini longitesta, H. d. dujardini oraz H. d. edlaueri. Podstawą wydzielenia dwóch pierwszych podgatunków była większa wysmukłość muszli(W/H) i wyższa względna wysokość ostatniego zwoju (Hlw/H) H. d. longitesta. Koncepcję tę zaakceptował Kókay (1966), ale i w tym przypadku okazy zilustrowane przez oboje autorów mieszczą się w unimodalnym spektrum zmienności okazów korytnickich. Cechą diagnostyczną H. d. edlaueri mają być szczególnie wydatne osiowe fałdy na ostatnim zwoju, ale osiowe fałdy wykazują gradację wydatności w ramach zmienności korytnickiej populacji S. schoenni. Wątpliwości, co do zasadności wydzielenia tych podgatunków potwierdza opis Strausza (1966), który zaznaczył, iż w badanym przez niego materiale można wprawdzie wyróżnić podgatunki wydzielone przez Beer-Bistricky, lecz okazów przejściowych jest znacznie więcej!
Wnioski
Z powyższych przykładów można wyciągnąć następujące wnioski:
1. Typologiczne podejście do zagadnienia gatunku prowadzi z reguły do wyodrębniania nadmiernej liczby jednostek systematycznych (gatunków, podgatunków, odmian), zaś przyjęcie koncepcji populacyjnej redukuje liczbę nowych taksonów.
2. Pracując zgodnie z populacyjną koncepcją gatunku należy z krytycyzmem podchodzić do danych literaturowych, gdyż mogą one reprezentować stanowisko typologiczne.
3. Poprawne wydzielanie gatunków kopalnych wymaga znajomości ontogenezy i zakresu zmienności na poszczególnych stadiach rozwojowych. Warunki te nie zawsze jednak mogą być spełnione.
Miarą trafności podejmowanych przez paleontologów decyzji taksonomicznych jest to, czy wydzielane przez nich gatunki kopalne odpowiadają realnie istniejącym niegdyś gatunkom biologicznym. Ujęcie populacyjne pozwala na spełnienie tego postulatu. Zmienność wewnątrzgatunkowa (wewnątrz- i międzypopulacyjna) jest bowiem immanentną właściwością żywych organizmów, zaś jej rozkład ma zwykle charakter normalny. Powszechność takiego rozkładu można zrozumieć wyłącznie poprzez pryzmat genetyki i paradygmatu darwinowskiego - jest bowiem efektem wymiany genów w procesach płciowych i działania doboru stabilizującego.
Literatura
Beer-Bistricky, E. 1958. Die miozänen Buccinidae und Nassaridae des Wiener Beckens und Niederösterreischs. Mitteilungen der Geologisches Gesllschaft Wien 49, 41-84.
Boettger, O. 1906. Zur Kenntnis der Fauna der mittelmiozänen Schichten von Kostej im Krasso-Szorenyer Komitat. Verhandlungen und Mitteilungen der Siebenburgisches Verein der Naturwisenschaften 54, 1-99.
Hilber, V. 1879. Neue Conchylien aus den mittelsteierischen Mediterranschichten. Sitzungberichte der Akademie der Wissenschafter, Mathematisch-Naturwissenschafliche Klasse 79, 416-464.
Hoernes, R. & Auinger, M. 1882. Die Gasteropoden der Meeres-Ablagerungen der erste und zweite miozänen Mediterran-Stufe in der Österreichisch-ungarischen Monarchie. Abhandlungen der Geologisches Reichsanstalt 12, 1-382.
Kókay, J. 1966. Geologische und paläontologische Untersuchung des Braunkohlengiebietes von Herend-Márkó (Bakony-Gebirge, Ungary). Geologia Hungarica, Series Palaeontologica 36, 1-147.
Strausz, L. 1966. Die miozän-mediterranen Gastropoden Ungarns, 1-692. Akadiemiai Kiadó. Budapest.
Venzo, S. & Pelosio, 1963. La malacofauna tortoniana del Colle di Vigoleno (Preappennino, Piacentino). Paleontographia Italiana 58, 43-113.
Dane genetyczne a kryteria morfologiczne gatunku otwornic planktonowych
WOJCIECH MAJEWSKI
Instytut Paleobiologii PAN
Problemy z systematyką otwornic od dawna pojawiały się przy wydzielaniu jednostek taksonomicznych różnego szczebla. Pierwsze rozbieżności dotyczące koncepcji gatunku w obrębie Foraminiferida doprowadziły już w połowie XIX wieku do ukształtowaniae odrębnych szkół taksonomicznych: brytyjskiej (lumpers) i kontynentalnej (splitters). Wiek XX przyniósł wzrost zainteresowania otwornicami, zwłaszcza planktonowymi, ze względu na ich przydatność w badaniach przede wszystkim stratygraficznych, ale także paleośrodowiskowych i ewolucyjnych. Doprowadziło to do sytuacji, gdy badania otwornic kopalnych i współczesnych zostały zdominowane przez geologów, a podziały taksonomiczne również form dzisiejszych zostały dokonane według koncepcji paleontologicznych. Podziały te opierają się wyłącznie na morfologii skorupek i jak się dziś okazuje doprowadziły do znaczącego zaniżenia oceny różnorodności gatunkowej tej grupy organizmów, przynajmniej w przypadku form dzisiejszych (Norris & de Vargas 2000).
Ostatnie dziesięciolecie przyniosło pierwsze badania materiału genetycznego dzisiejszych otwornic. Do celów badawczych wykorzystano sekwencje konserwatywnego ewolucyjnie genu małej podjednostki rybosomu (SSU rDNA), który dobrze odzwierciedla stosunki filogenetyczne. Ostatnio przeprowadza się także badania regionu ITS rDNA, który podlega ~5 razy częstszej mutacji niż SSU rDNA (de Vargas et al. 2001), co umożliwia dokładne śledzenie różnorodności genetycznej nawet w obrębie poszczególnych populacji. Już pierwsze wyniki badań molekularnych wykazały, że otwornice jako grupa należą do najszybciej ewoluujących organizmów, a otwornice planktonowe mają ~100 razy wyższe tempo mutacji niż otwornice bentosowe (de Vargas et al. 1997).
Wiele gatunków morfologicznych dzisiejszych otwornic planktonowych to grupy odmiennych genotypów. Każdy z nich wypada uznać za samodzielny gatunek biologiczny ze względu na różnice genetyczne, a także ekologiczne i biologiczne dowodzące izolacji genetycznej populacji, które reprezentują (Huber et al. 1997; de Vargas et al. 1999, 2001). Znamienne, że wcześnie rozpoznane wyraźne różnice morfologiczne miedzy genotypami, np. lewo- lub prawo-skrętność u Neogloboquadrina pachyderma (Ericson 1959), czy różnice w porowatości i średnicy skorupek Orbulina universa (Bé 1973), były powszechnie interpretowane jako naturalna różnorodność w obrębie populacji lub zmienność ontogenetyczna wywołana warunkami środowiska.
Huber i inni (1997) wykazali obecność dwóch odrębnych genotypów w obrębie pojedynczego gatunku morfologicznego Globigerinella siphonifera. Już wcześniej rozpoznano dwa typy tej otwornicy różniące się budową i kształtem pseudopodiów, pigmentacją, rodzajem symbiotycznych glonów, zachowaniem w laboratorium, czy wartościami izotopowymi węgla i tlenu w ich skorupkach. Z drugiej jednak strony oba typy G. siphonifera nie wykazywały jednoznacznej odmienności morfologii skorupek (poza różnicami w porowatości), co było podstawą do zaliczenia ich do jednego gatunku morfologicznego. Tymczasem różnice sekwencji SSU rDNA pomiędzy oboma typami są na tyle jednoznaczne i istotne, że wskazują na izolację genetyczna czyli obecność dwóch oddzielnych gatunków biologicznych. Analogiczna sytuacja istnieje w przypadku Orbulina universa (de Vargas et al. 1999) i Globorotalia truncatulinoides (de Vargas et al. 2001), w obrębie których wydzielono odpowiednio 3 i 4 genotypy (gatunki biologiczne) różniące się jedynie trzeciorzędnymi cechami morfologicznymi skorupki, ale za to mające odmienne wymagania ekologiczne. Badania genetyczne wykazały też obecność co najmniej dwóch genotypów (gatunków biologicznych) w obrębie Globigerina bulloides, Turborotalita quinqueloba, Neogloboquadrina pachyderma i Globigerinoides ruber (Stewart et al. 2001; Darling et al. 1999, 2000).
Obok wyraźnego podważenia paleontologicznej (morfologicznej) koncepcji gatunku dzisiejszych otwornic planktonowych, badania genetyczne wykazały zgodność ich filogenezy z klasyfikacją taksonomiczną na poziomie rodziny (de Vargas et al. 1997, de Vargas & Pawłowski 1998). Określenie stopnia odmienności genetycznej i tempa mutacji w obrębie poszczególnych linii filogenetycznych daje nadzieję na weryfikację zasadności wyróżnienia rodzajów, czego przykładem jest casus rodzaju Globigerinella (Huber et al. 1997).
Literatura
Bé, A.W.H. 1973. Orbulina universa d'Orbigny in the Indian Ocean. Micropaleontology 19, 150-192.
Darling, K.F., Wade, C.M., Kroon, D., Brown, A.J.L., & Bijma, J. 1999. The diversity and distribution of modern planktonic foraminiferal small subunit ribosomal RNA genotypes and their potential as tracers of present and past ocean circulation. Paleoceanography 14, 3-12.
Darling, K.F., Wade, C.M., Stewart, I.A., Kroon, D., Dingle, R. & Brown, A.J.L. 2000. Molecular evidence for genetic mixing of Arctic and Antarctic subpolar populations of planktonic foraminifers. Nature 405, 43-47.
de Vargas, C., Norris, R., Zaninetti, L., Gibb, S.W. & Pawłowski, J. 1999. Molecular evidence of cryptic speciation in planktonic foraminifers and their relation to oceanic provinces. Proceedings of the National Academy of Sciences USA 96, 2864-2868.
de Vargas, C. & Pawłowski, J. 1998. Molecular versus taxonomic rates of evolution in planktonic foraminifera.- Molecular Phylogenetics and Evolution 9, 463-469.
de Vargas, C., Renaud, S., Hilbrecht, H. & Pawłowski, J. 2001. Pleistocene adaptive radiation in Globorotalia truncatulinoides: genetic, morphologic, and environmental evidence. Paleobiology 27, 104-125.
de Vargas, C., Zaninetti, L., Hilbrecht, H. & Pawłowski, J. 1997. Phylogeny and rates of molecular evolution of planktonic foraminifera: SSU rDNA sequences compared to the fossil record. Molecular Evolution 45, 285-294.
Ericson, F.B. 1959. Coiling direction of Neogloboquadrina pachyderma as a climatic index. Science 130, 219-220.
Huber, B.T., Bijma, J. & Darling, K. 1997. Cryptic speciation in the living planktonic foraminifer Globigerinella siphonifera (d'Orbigny). Paleobiology 23, 33-62.
Norris, R.D. & de Vargas, C. 2000. Evolution all at see. Nature 405, 23-24.
Stewart, I.A., Darling, K.F., Kroon, D., Wade, C.M., & Troelstra, S.R. 2001. Genotypic variation in subarctic Atlantic planktic foraminifera. Marine Micropaleontology 43, 143-153.
Egzekwowanie zasady priorytetu w taksonomii paleozoologicznej
JERZY DZIK
Instytut Paleobiologii PAN
Szkielet znaczącej części kopalnych organizmów składa się z oddzielnych elementów, które przechodzą osobno do osadu po rozkładzie tkanek miękkich i osobno są wydobywane ze skały. Niekiedy różnią się składem mineralnym, albo wręcz zachowuja się w całkiem odmienny sposób. Zwykle przyjmuje się to za oczywiste i żadnych szczególnych uzasadnień nie wymaga łączenie ze sobą w jednym gatunku tarczy głowowych i ogonowych trylobitów czy różnych kości szkieletu kręgowców. Wątpliwości pojawiają się, kiedy różne części jednego zwierzęcia rzadko współwystępują ze sobą (np. muszle i aptychy amonitów), albo też powiązanie ich ze sobą jest trudne, bo niełatwo rozdzielić podobne skamieniałości należące do licznych współwystępujących gatunków. Tak jest w badaniach szczątków roślin naczyniowych, aparatów gębowych pierścienic (skolekodontów), czy konodontów. Przez długi czas nie było odmienności w podejściu do tego zagadnienia na gruncie paleobotaniki i paleobiologii. W ostatnich dekadach ubiegłego wieku doszło jednak do przełomu metodologicznego w badaniach konodontów, który rozszerzył się na inne dziedziny paleozoologii. Analogiczne tendencje zaznaczają się i w paleobotanice. To jest powód, dla którego warto się zapoznać z metodologicznymi podstawami postępowania w takich sytuacjach.
Zagadnienie to ma dwa osobne aspekty: (1) metodę postępowania, która pozwala na odtworzenie składu kompletnego zestawu elementów szkieletowych, czy wręcz części ciała osobników jednego gatunku oraz (2) sposób wyselekcjonowania właściwej nazwy gatunkowej i rodzajowej z dostępnej listy synonimów. Metodyka wiązania ze sobą skamieniałości należących do jednego organizmu została szczegółowo rozpracowana na przykładzie konodontów. Nie jest to bezpośrednio związane z tematyką kursu, napomknę więc jedynie, że opiera się o zasadę statystycznego współwystępowania ze sobą elementów jednego gatunku i możliwość odnalezienia takich kombinacji występowania gatunków, że możliwe jest wykluczenie którejś z równoprawnych roboczych rekonstrukcji zestawu. Pomocne jest morfologiczne podobieństwo różnych części należących do jednego gatunku i możliwość homologizacji części zestawów należących do różnych gatunków. Pozwala to na testowanie hipotez na podstawie założenia, że wszystkie stadia ewolucji w jednym ciągu gatunków winny mieć w pełni homologiczne zestawy elementów. W szczególności skuteczne okazało się podejście polegające na formułowaniu hipotez o składzie zestawów i testowanie ich przez nowe znaleziska. W ciągu ćwierćwiecza rozpoznany został skład aparatów gębowych większości konodontów, od kambru do końca triasu.
Metoda wyznaczania nazwy zrekonstruowanego jakimkolwiek sposobem i nie dającego się obalić wyobrażenia o składzie zestawu części ciała organizmu kopalnego opiera się wyłącznie o zasadę egzekwowania priorytetu. W gruncie rzeczy jest to wybór właściwego holotypu. Wymaga to najpierw określenia granic gatunku. Jesteśmy na gruncie paleontologii, chodzi więc o gatunek temporalny (chronospecies), czyli serię populacji obejmujących obiektywnie określoną przestrzeń geograficzną i arbitralnie wyznaczony wycinek czasu geologicznego. Wśród populacji tych jest zwykle pewna liczba populacji typowych wcześniej nazwanych gatunków. Z reguły wprowadzonych na podstawie pojedynczych elementów szkieletu, przed rekonstrukcją pełnego (czy choćby częściowego) ich zestawu. Wzorcem każdego z tych gatunków jest holotyp. Jego locus typicus określa położenie w przestrzeni geograficznej, stratum typicum w czasie. Wystarcza więc zebrać wszystkie holotypy, których loci typici i strata typica są w obrębie rekonstruowanego gatunku. Holotyp, który ma najstarszą datę wprowadzenia jest jego holotypem, a nazwa gatunkowa nazwą do niego przywiązaną. Wszystkie inne są synonimami. Do konkurencji nie wchodzą nazwy, których holotypy są poza zasięgiem gatunku, choćby nawet kiedyś były użyte do określenia którejś z części ciała teraz włączanej do rekonstrukcji. Nie ma więc w tym przypadku żadnego sensu zastanawianie się nad listami synonimów z pominięciem ich typów opisowych.
Na podobnych zasadach oparty jest wybór nazwy rodzajowej. Tu miejsce holotypu zajmuje gatunek typowy. Określić trzeba najpierw zakres rodzaju, czyli wyliczyć wszystkie włączane doń gatunki temporalne. Wśród synonimów tych gatunków są gatunki typowe rodzajów. Jeśli ich holotypy znajdą się w obrębie rodzaju, ich gatunki konkurują o godność jego gatunku typowego. Nierzadko gatunkiem typowym okazuje się młodszy synonim jednego z gatunków zaliczonych do rodzaju.
Istotnym aspektem tego rozumowania jest to, że abstrahuje od wartości diagnostycznej holotypów. Ważne jest tylko to, by z pewną wiarygodnością można było przyjąć, że należy do określonej populacji. Oznacza to, że rozważania dotyczą wyłącznie locus typicus i stratum typicum, czyli w istocie stanowiska kopalnego bądź próbki, do której należy holotyp. Jeśli zawiera szczątki tylko jednego gatunku, wątpliwości, że holotyp do niego należy być nie może, choćby był pozbawiony jakichkolwiek cech charakterystycznych. Jeśli gatunków w próbce jest więcej, trzeba zdecydować, do którego z nich holotyp należy w oparciu o informacje morfologiczne, ale w ostateczności wybór może być arbitralny. To prawo pierwszego rewidenta, czyli badacza, który dokonał rekonstrukcji. To on decyduje, jaki nadać sens wątpliwemu holotypowi. Brzmi to nieprzyjemnie, ale alternatywa byłaby jeszcze gorsza. Poglądy o tym, które cechy są ważne, a które nie, zmieniają się w trakcie rozwoju nauki; różnie też do tego podchodzą poszczególni uczeni. Zasada decyzji pierwszego rewidenta jest zgodna z duchem kodeksów nomenklatury i ucina zbyteczne i niekończące się zwykle spory nomenklatoryczne. Jest gwarancją stabilności nomenklatorycznej, a to jest z kolei podstawą reputacji taksonomii. Gorszące spory o zagadnienia nie mające żadnego znaczenia naukowego dostatecznie ją już osłabiły.
Literatura
Bengtson, S. 1985. Taxonomy of disarticulated fossils. Journal of Paleontology 59, 1350-1358.
Boczarowski, A. 2001. Isolated sclerites of Devonian non-pelmatozoan echinoderms. Palaeontologia Polonica 59, 1-219.
Boogaard, M. van den & Kuhry, M.J.M. 1979. Statistical reconstructionnof the Palmatolepis apparatus (Late Devonian conodontophorids) at the generic, subgeneric, and specific level. Scripta Geologica 49, 1-57.
Dzik, J. 1991. Evolution of oral apparatuses in the conodont chordates. Acta Palaeontologica Polonica 36, 265-323.
Pisera, A. & Dzik, J. 1979. Tithonian crinoids from Rogoźnik (Pieniny Klippen Belt, Poland) and their evolutionary relationships. Eclogae geologicae Helvetiae 72, 805-849.
|