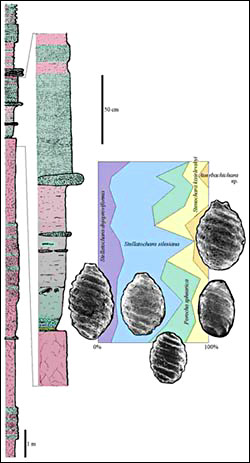
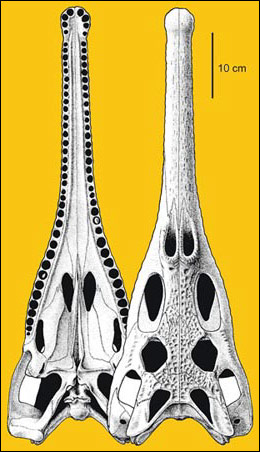
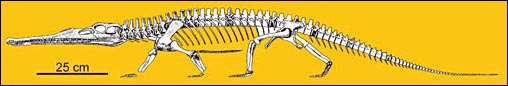
Aetozaury. Aetozaury, rodzina Stagonolepididae, to stosunkowo
niewielkie, ponadmetrowej długości, tekodonty z rozbudowanym pancerzem z płyt
kostnych wokół ciała. Ich pyski wydłużone były na końcu w niedługi ryj, a tępe
zęby były odmiennych rozmiarów na kościach szczękowej i przedszczękowej, co
sugeruje przystosowanie do roślinnego pokarmu. W materiałach z Krasiejowa mamy
połamaną na kilka części czaszkę aetozaura, 11 płyt kostnych i zapewne kości
długie, których nie potrafimy jeszcze odróżnić od kości fitozaurów. Nie
potrafimy jescze zidentyfikować rodzaju, choć radialne urzeźbienie, wyrazisty
guz z tyłu i gładki margines wzdłuż przedniej krawędzi wykluczają identyczność z
większością form znanych z literatury. Wraz z Paleorhinus w amerykańskich
stanowiskach występuje aetozaur Longosuchus, którego płyty (Hunt &
Lucas 1990; Long & Murry 1995) wyraźnie różnią się od krasiejowskich
mniejszą wypukłością i wyższym guzem, oraz Coahomasuchus, o płytach bez
radialnej ornamentacji (Heckert & Lucas 1999). Śląskie aetozaury były od
nich mniej wyspecjalizowane ewolucyjnie, co jest kolejną sugestią na rzecz
znacznego wieku geologicznego warstw z Krasiejowa.
Dinozaury. Wśród kości z Krasiejowa szczątki ściśle lądowych gadów są
rzadkie. Wśród nich szczególnie intrygujący jest pojedynczy tylny kręg krzyżowy
ZPAL AbIII/284 o budowie odmiennej od tekodontów i fragmenty lewych kości
szczękowych ZPAL AbIII/281i 282 z zachowanym po jednym zębie w zębodołach.
Szczególną cechą kręgu krzyżowego jest pionowe rozszerzenie wyrostka
poprzecznego wraz z żebrem i płaska przednia powierzchnia trzonu, dowodząca
ścisłego powiązania (ale nie zrośnięcia) kręgów w obrębie "kości krzyżowej"
(kręgi krzyżowe fitozaurów mają powierzchnie trzonów takie, jak pozostałe
kręgi). Kręg ten jest bardzo podobny do odpowiedniego kręgu
Staurikosaurus z późnego triasu Brazylii (Galton 1999: fig. 8D-F) i
Chindeosaurus z formacji Chinle Arizony (Long & Murry 1995; Hunt
et al. 1998), przedstawicieli herrerasauridów - najpierwotniejszych
drapieżnych dinozaurów. Znaleziska amerykańskie są późniejsze od doby
Paleorhinus.
Z kolei ząb, którego wierzchołek widoczny jest wewnątrz zębodołu kości
zębowej ZPAL AbIII/282, różni się od podobnego kształtu i rozmiarów zębów
policzkowych fitozaurów orientacją poszczególnych ząbków piłkowania krawędzi. U
fitozaurów, jak to dobrze widać na krasiejowskich zębach Paleorhinus,
ząbki piłki ustawione są prostopadle do krawędzi ostrza. Ząbki piłki tego zęba
są zaś ustawione równolegle do jego wierzchołka. Przypomina to sytuację typową
dla roślinożernych dinozaurów, choć gęstość piłkowania jest w tym przypadku
porównywalna do typowej dla fitozaurów. Również nieregularne podłużne żebra na
powierzchni tego zęba nadają mu postać odmienną od gładkich zębów fitozaurów.
Niekompletna kość szczękowa ZPAL AbIII/281 nie jest podobna do odpowiednich
kości drapieżnych herrerasauridów, jest natomiast porównywalna z kośćmi
wczesnych dinozaurów ptasiomiednicznych Lesothosaurus i
Pisanosaurus.
Dostępny materiał jest zbyt ubogi, by stwierdzić, czy kręg i fragment szczęki
należały do tego samego gatunku. Zwierzęta te były stosunkowo niewielkich
rozmiarów (około metra długości), stopień skostnienia kręgu dowodzi jednak, że
mamy do czynienia z osobnikiem dorosłym. Bez wątpienia, jako najstarsze
wystąpienie kostnych szczątków dinozaurów w Europie, jest to znalezisko o
wielkim znaczeniu naukowym. Można mieć nadziejEA^, że przyszłe wykopaliska
dostarczą dodatkowego materiału szkieletowego (w szczególności diagnostycznych
dla dinozaurów kości miednicy), który pozwoli na precyzyjniejsze określenie
pokrewieństw tego zwierzęcia.
Podziękowania
Podczas prac wykopaliskowych na terenie wyrobiska kopalni Krasiejów
korzystaliśmy z pomocy technicznej ze strony Cementowni Strzelce Opolskie S.A.
Składamy za to serdeczne podziękowania jej dyrekcji, a w szczególności
Dyrektorowi do Spraw Technicznych, panu mgr inż. Ernestowi Jelito. Podczas
opracowywania materiałów bardzo pomocne były dla nas merytoryczne sugestie pani
doc. Magdaleny Borsuk-Białynickiej i pani prof. Halszki Osmólskiej.
Literatura
Arcucci, A. & Marsicano, C.A.1998. A distinctive new archosaur from the
Middle Triassic (Los Chanares Formation) of Argentina. Journal of Vertebrate
Paleontology 18, 228-232.
Ballew, K.L. 1989. A phylogenetic analysis of Phytosauria from the later
Triassic of the western United States. In S. G. Lucas & A. P.
Hunt (eds) Dawn of the Age of Dinosaurs in the American Southwest,
309-339, New Mexico Museum of Natural History, Albuquerque.
Benton, M.J. 1989. Mass extinction among tetrapods and the quality of the
fossil record. Philosophical Transactions of the Royal Society of
London B325, 369-386.
Bilan W. 1975. Profil retyku w Krasiejowie koło Opola. Zeszyty Akademii
Górniczo-Hutniczej, Geologia 1 (3), 13-20.
Borsuk-Białynicka, M., Cook, E., Evans, S.E., & Maryańska, T. 1999. A
microvertebrate assemblage from the Early Triassic of Poland. Acta
Palaeontologica Polonica 44, 167-188.
Buffetaut, E. & Ingavat, R. 1982. Phytosaur remains (Reptilia,
Thecodontia) from the upper Triassic of north-eastern Thailand. Geobios
1, 7-17.
Buffetaut, E. 1993. Phytosaurs in time and space. Paleontologia Lombarda
della Societa Italiana di Science Naturali e del Museo Civico di Storia
Naturale di Milano, Nuova Serie 2, 39-44.
Chatterjee, S. 1978. A primitive parasuchid (phytosaur) reptile from the
upper Triassic Maleri Formation of India. Palaeontology
21, 83-127.
Chowdhury, T. R. 1965. A new metoposauroid amphibian from the Upper Triassic
Maleri formation of Central India. Philosophical Transactions of the Royal
Society London B250, 1-52.
Colbert, E.H. & Imbrie, J. 1956. Triassic metoposaurid amphibians.
Bulletins of the American Museum of Natural History 110,
399-452.
Deczkowski, Z, Marcinkiewicz, T, & Maliszewska A. 1997. Noryk i retyk.
In: S. Marek & M. Pajchlowa (eds) Epikontynentalny perm i mezozoik w Polsce.
Prace Państwowego Instytutu Geologicznego 153,174-194.
Dutuit, J. M. 1976. Introduction a l'etude paleontologique du Trias
continental marocain. Description des premiers Stegocephales recueillis dans le
couloird'Argana (Atlas occidental). Memoires du Museum National
d'Histoire Naturelle, Paris, Ser. C 36: 1-253.
Dutuit, J.M. 1977a. Description du crane de Angistorhinus talainti
nov. sp., Un nouveau phytosaure du Trias atlasique marocain. Bulletin de
la Museum National de Histoire Naturelle de Paris, 3e ser. 489,
297-337.
Dutuit, J.M. 1977b. Paleorhinus magnoculus, phytosaure du Trias
superieur de l'Atlas marocain. Geologie Mediterranee 4,
255-268.
Dutuit, J. M. 1978. Description de quelques fragments osseux provenant de la
region de Folakara (Trias superieur malgache). Bulletin de la Museum National
de Histoire Naturelle de Paris, 3e ser. 516, 79-89.
Fowell, S.J., Cornet, B., & Olsen, P.E. 1994. Geologically rapid Late
Triassic extinxtions: Palynological evidence from the Newark Supergroup.
Geological Society of America Special Paper 288, 197-206.
Fraas, E. 1913. Neue Labiryntodonten der Schwäbischen Trias.
Palaeontographica 60, 275-294.
Gajewska, I. 1973. Charakterystyka osadów piaskowca trzcinowego na Niżu
Polskim. Kwartalnik geologiczny 17, 507-515.
Galton, P.M. 1999. Sex, sacra and Sellosaurus gracilis (Saurischia,
Sauropodomorpha, Upper Triassic, Germany) or why the character "two sacral
vertebrae" is plesiomorphic for Dinosauria. Neues Jahrbuch für
Geologie und Paläontologie, Abhandlungen 213,
19-55.
Gans, C.L. & Tinkle, D. 1977. Ecology and Behavior. Biology of
Reptilia 7, 229-235. Academic Press, London.
Godefroit, P. & Cuny, G. 1997. Archosauriform teeth from the Upper
Triassic of Saint-Nicolas-de-Port (northeastern France). Palaeovertebrata
26, 1-34.
Gregory, J.T. 1962. The genera of phytosaurs. American Journal of Science
260, 652-690.
Gregory, J.T. 1969. Evolution und interkontinentale Beziehungen der
Phytosauria (Reptilia). Paläontologische Zeitschrift
43, 37-51.
Gregory, J. T. 1980. The otic notch of metoposaur labirynhodonts. In: L.L.
Jacobs (eds.) Aspects of Vertebrate History. 125-136. Museum of
Northern Arizona Press Flagstaff.
Gregory, J.T. & Westphal, F. 1969. Remarks on the phytosaur genera of the
European Trias. Journal of Paleontology 43, 1296-1298.
Grodzicka-Szymanko, W. & Orłowska-Zwolińska, T. 1972. Stratygrafia
górnego triasu NE części obrzeżenia Górnośląskiego Zagłębia Węglowego.
Kwartalnik Geologiczny 16, 216-232.
Gryglas, E. 1999. Ramienice a środowisko życia fytozaurów i metopozaurów z
późnego triasu Śląska Opolskiego. 72 pp. Praca magisterska wykonana w
Zakładzie Paleontologii Uniwersytetu Warszawskiego.
Heckert, A.B. & Lucas, S.G. 1999. A new aetosaur (Reptilia: Archosauria)
from the Upper Triassic of Texas and the phylogeny of aetosaurs. Journal of
Vertebrate Paleontology 19, 50-68.
Huber, P., Lucas, S.G., & Hunt, A. P. 1993. Vertebrate Biochronology of
the Newark Supergroup, Triassic, Eastern North America. In S.G. Lucas, & M.
Morales (eds.) The Nonmarine Triassic. New Mexico Museum of Natural
History & Science Bulletin 3, 179-186 .
Huene, F. Von, 1939. Ein primitiver Phytosaurier in der jüngeren
nordostalpinen Trias. Zentralblatt der Mineralogie, Geologie und
Paläontologie 4, 139-144.
Hunt, A.P. 1993. Revision of the Metoposauridae (Amphibia: Temnospondyli) and
description of a new genus from western north America. In: M. Morales (eds)
Aspects of Mesozoic Geology and Paleontology of the Colorado Plateau.
Museum of northern Arizona bulletin 59, 67-97.
Hunt, A. P. & Lucas, S. G. 1990. Re-evaluation of "Typothorax"
meadai, a Late Triassic aetosaur from the United States.
Paläontologische Zeitschrift 64, 317-328.
Hunt, A. P. & Lucas, S. G. 1991. The Paleorhinus biochron and the
correlation of the non-marine upper triassic of Pangea. Palaeontology
34: 478-501.
Hunt, A. P., Lucas, S. G., Heckert, A.B., Sullivan, R.M., & Lockley, M.G.
1998. Late Triassic dinosaurs from the western United States. Geobios
31, 511-531.
Jaekel, O. 1910. Über einen neuen Belodonten aus dem Buntsandstein von
Bernburg. Sitzungsbericht der Gesellschaft naturforschender Freunde zu
Berlin 5, 197-229.
Kirby, R. E. 1993. Relationship of Late Triassic basin evolution and faunal
replacement events in the southwestern United States: Perspectives from the
upper part of the Chinle Formation in Northern Arizona. In: S.G. Lucas, & M.
Morales (eds) The Nonmarine Triassic. New Mexico Museum of Natural History
& Science Bulletin 3, 233-242.
Kłapciński, J. 1993. Litostratygrafia profili głębokich otworów wiertniczych
w regionie opolskim. Acta Universitatis Wratislaviensis 8,
1-129.
Kotański, Z. 1996. History of discovery and age of labyrinthodont remains in
the Tatra Mts, Poland. Prace Muzeum Ziemi 43, 47-52.
Kotlicki, S. & A. Kubicz, A. 1974. Trias Śląska Opolskiego. In: J.
Rutkowski (ed.) Przewodnik XLVI Zjazdu Polskiego Towarzystwa
Geologicznego, Opole 12-14 września 1974. 18-26 Wydawnictwa Geologiczne,
Warszawa.
Kuhn, O. 1933. Labyrinthodonten und Parasuchier aus dem mittleren Keuper von
Ebrach in Oberfranken. Neues Jahrbuch für Mineraliogie,
Geologie und Paläontologie, Abteilung B, Beilageband
69, 94-143.
Kuhn, O. 1936. Weitere Parasuchier und Labyrinthodonten aus dem
Blasensandstein des mittleren Keuper von Ebrach. Palaeontographica
A86, 61-98.
Kunisch, H. 1890. Labirynthodontenreste des oberschlesischen Muschelkalks.
Zeitschrift der Deutschen Geologischen Gesellschaft 42,
377-385.
Long, R.A. & Murry, P.A. 1995. Late Triassic (Carnian and Norian)
tetrapods from the southwestern United States. New Mexico Museum of Natural
History & Science Bulletin 4, 1-254.
Lucas, S.G. 1998. Global Triassic tetrapod biostratigraphy and biochronology.
Palaeogeography, Palaeoclimatology, Palaeoecology 143,
347-384.
Mader, D. 1997. Palaeoenvironmental Evolution and Bibliography of the
Keuper (Upper Triassic) in Germany, Poland and other parts of Europe.
1058 pp. Sven von Loga Verlag, Köln.
Maliszewska, A. 1972. Geneza brekcji lisowskiej na podstawie badań
petrograficznych. Biuletyn Instytutu Geologicznego 261,
33-54.
Maryańska, T. & Shishkin, M.A. 1996. New cyclotosaurid (Amphibia:
Temnospondyli) from the Middle Triassic of Poland and some problems of
interrelationships of capitosauroids. Prace Muzeum Ziemi 43,
53-83.
Milner, A.R. 1990. The radiation of temnospondyl amphibians. In: P.D. Taylor
& G.P. Larwood (eds) Major Ewolutionary Radiations. 321-349.
Clarendon Press, Oxford.
Milner, A.R. 1994. Late Triassic and Jurassic ampibians: fossil record and
phylogeny. In N.C. Fraser, & H.D. Sues, (eds.) In the Shadow of
Dinosaurs, Early Mesozoic Tetrapods. 5-22. Cambridge University Press,
Cambridge.
Murry, P.A. 1986. Vertebrate paleontology of the Dockum Group, western Texas
and eastern New Mexico In K. Padian, (eds.) The Begining of the Age of the
Dinosaurs: Faunal Changes Between Triassic-Jurassic Boundary. 109-137.
Cambridge University Press, Cambridge.
Mutti M. & Weissert H., 1995.Triassic monsoonal climate and its signature
in Ladinian - Carnian carbonate platforms (Southern Alps, Italy). Journal of
Sedimentary Research B65, 357 - 367.
Orłowska-Zwolińska, T. 1983. Palinostratygrafia epikontynentalnych osadów
wyższego triasu w Polsce. Prace Instytutu Geologicznego
104, 1-88.
Padian, K. 1989. Did > survive the Triassic? In S. G. Lucas &
A. P. Hunt (eds) Dawn of the Age of Dinosaurs in the American
Southwest, 401-414, New Mexico Museum of Natural History,
Albuquerque.
Parrish, J. M. 1989. Late Triassic tetrapods of the north American southwest.
In S. G. Lucas & A. P. Hunt (eds) Dawn of the Age of Dinosaurs in
the American Southwest, 401-414, New Mexico Museum of Natural History,
Albuquerque.
Pieńkowski, G. 1988. Analiza facjalna najwyższego triasu i liasu Wyżyny
Krakowsko-Wieluńskiej oraz perspektywy występowania surowców ilastych.
Przegląd geologiczny 36, 449-456.
Renesto, S. & Lombardo, C. 1999. Strusture of the tail of a phytosaur
(Reptilia, Archosauria) from the Norian (Late Triassic) of Lombardy (Northern
Italy). Rivista Italiana di Paleontologia e Stratigrafia 105,
135-144.
Renesto, S. & Paganoni, A. 1998. A phytosaur skull from the Norian (Late
Triassic) of Lombardy (Northern Italy). Rivista Italiana di Paleontologia e
Stratigrafia 104, 115-122.
Roemer, F. 1870. Geologie von Oberschlesien. 587 pp. Nischkowsky,
Breslau.
Rogers, R., Swisher, C., Monetta, A., Forster, C, & Martinez, R. 1993.
The Ischigualasto tetrapod assemblage (Late Triassic, Argentina) and
40Ar/39Ar dating on dinosaur origin. Science
260, 794-797.
Simms, M.J., Ruffell, A.H., & Johnson, A.I.A. 1995. Biotic and climatic
changes in the Carnian (Triassic) of Europe and adjacent areas. In: N.C. Fraser,
& H.D. Sues (eds.), In the Shadow of Dinosaur, Early Mesozoic
Tetrapods. 5-22. Cambridge University Press, Cambridge.
Volz, W. 1902. Proneusticosaurus, eine neue Sauropterygier-Gattung aus
dem unteren Muschelkalk Oberschlesiens. Palaeontographica 49,
121-162.
Walker, A.D. 1968. Protosuchus, Proterochampsa and the origin of
phytosaurs and crocodiles. Geological Magazine 105, 1-14.
Warren, A. & Snell, N. 1991. The postcranial skeleton of Mesozoic
temnospondyl amphibians: a review. Alcheringa 15, 43-64.
Westphal, F. 1970. Phytosaurier-Hautplatten aus der Trias von Madagascar -
ein Beitrag zur Gondwana-Paläogeographie. Neues Jahrbuch für
Geologie und Paläontologie, Monatshefte 1970,
632-638.
Westphal, F., 1976. Phytosauria. In O. Kuhn (ed.) Handbuch der
Paläoherpetologie 13, 99-120. Gustav Fischer Verlag,
Stuttgart.
Summary. A new Paleorhinus fauna has been discovered
in a grey claystone corresponding to the subsurface Drawno beds, which
are exposed in a cement plant claypit at Krasiejów near Opole in southern
Poland. The vertebrate assemblage of perfectly preserved skeletons,
isolated skulls, and postcranial remains is dominated with
labirynthodonts Metoposaurus diagnosticus, the phytosaur
Paleorhinus sp. being the second in number. Aetosaurs are
represented by an incomplete skull, scuta and fragmentary bones. One sacral bone
most probably representing an early dinosaur related to
Staurikosaurus or Chindesaurus and two incomplete maxilla with
teeth showing serration parallel to the cusp have also been found. The
Krasiejów assemblage includes also dipnoans Ceratodus, undetermined fish
scales and teeth, numerous unionid bivalves, very abundant characean
gyrogonites, and ostracods. The Paleorhinus species is relatively
short-snouted and may represent an earlier stage in the evolution of the
lineage than the populations from Blasensandstein in Franconia and other known
Paleorhinus faunas. This is consistent with the presence of
Metoposaurus diagnosticus, the species originally described from
the Schilfsandstein, located below the Blasensandstein. An equivalent of
Schilfsandstein is known in boreholes drilled nearby the locality and the
bone-bearing horizon is about 80 m above its top, and about 20 m above
the last occurrence of gypsum in the section. Lehrberg Schichten in
Germany seem thus to be its lateral equivalents, both presumably
representing the final event in the middle to late Carnian eustatic cycle. The
Paleorhinus fauna was apparently persistent in the area, as an
incomplete skeleton has been found about 8 m below the most fossiliferous
horizon, in a calcareous intercalation within red clays comparable with
the German Rote Wand.
* Zanclodon arenaceus podobno nie jest jednak fitozaurem.